Сталь ст2 характеристики: Сталь СТ2 по ГОСТ – характеристики, химический состав
alexxlab | 04.12.1988 | 0 | Разное
Ст2сп сталь: характеристики и расшифовка, применение и свойства стали
- Стали
- Стандарты
Всего сталей
| Страна | Стандарт | Описание | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Россия | ГОСТ 380-2005 |  Марки Марки | ||||||||||
| Россия | ГОСТ 535-2005 | Прокат сортовой и фасонный из стали углеродистой обыкновенного качества. Общие технические условия | ||||||||||
| Россия | ГОСТ 13663-86 | Трубы стальные профильные. Технические требования | ||||||||||
| Россия | ГОСТ 14637-89 | Прокат толстолистовой из углеродистой стали обыкновенного качества.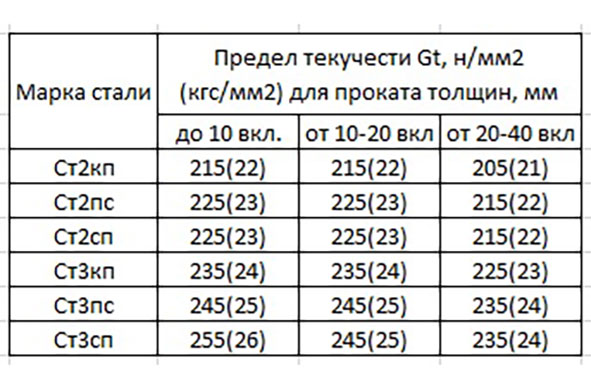 Технические условия Технические условия | ||||||||||
Механические свойства Ст2сп
Механические свойства по стандарту ГОСТ 380-2005
|
Предел текучести, σ0,2, МПа |
Временное сопротивление разрыву, σв, МПа |
Относительное удлинение при разрыве, δ5, % |
| 195 – 225 | 335 – 430 |
29 – 32 |
Механические свойства по стандарту ГОСТ 13663-86
|
Сортамент |
Тип трубы |
Обработка |
Предел текучести, σ0,2, МПа | Временное сопротивление разрыву, σв, МПа | Относительное удлинение при разрыве, δ5, % |
|
Труба |
Электросварная |
Термически обработанная |
> 206 |
> 333 |
> 24 |
|
Труба |
Электросварная |
После прокатного нагрева |
> 206 |
> 333 |
> 24 |
|
Труба |
Электросварная |
Без термической обработки |
– |
> 353 |
> 10 |
Механические свойства по стандарту ГОСТ 14637-89
|
Состояние поставки |
Категория |
Толщина, мм |
Временное сопротивление разрыву, σв, МПа | Относительное удлинение при разрыве, δ5, % | Изгиб до параллельности сторон | |
|
Прокат толстолистовой |
1 |
4 – 20 |
> 225 |
330 – 430 |
> 32 |
d = 1,5a |
| Прокат толстолистовой |
1 |
20 – 40 |
> 215 |
330 – 430 |
> 31 |
d = 2,5a |
| Прокат толстолистовой |
1 |
40 – 100 |
> 205 |
330 – 430 |
> 29 |
d = 2,5a |
| Прокат толстолистовой |
1 |
100 – 160 |
> 195 |
330 – 430 |
> 29 |
d = 2,5a |
| Прокат толстолистовой |
2 |
4 – 20 |
> 225 |
330 – 430 |
> 32 |
d = 1,5a |
| Прокат толстолистовой |
2 |
20 – 40 |
> 215 |
330 – 430 |
> 31 |
d = 2,5a |
| Прокат толстолистовой |
2 |
40 – 100 |
> 205 |
330 – 430 |
> 29 |
d = 2,5a |
| Прокат толстолистовой |
2 |
100 – 160 |
> 195 |
330 – 430 |
> 29 |
d = 2,5a |
×
Отмена Удалить
×
Выбрать тариф
×
Подтверждение удаления
Отмена Удалить
×
Выбор региона будет сброшен
Отмена
×
×
Оставить заявку
×
| Название | |||
Отмена
×
К сожалению, данная функция доступна только на платном тарифе
Выбрать тариф
Ст2пс сталь: характеристики и расшифовка, применение и свойства стали
- Стали
- Стандарты
Всего сталей
| Страна | Стандарт | Описание | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Россия | ГОСТ 380-2005 | Сталь углеродистая обыкновенного качества. Марки | ||||||||||
| Россия | ГОСТ 535-2005 | Прокат сортовой и фасонный из стали углеродистой обыкновенного качества. Общие технические условия | ||||||||||
| Россия | ГОСТ 30136-95 | Катанка из углеродистой стали обыкновенного качества. Технические условия Технические условия | ||||||||||
 js_elem_306751″>
js_elem_306751″>Механические свойства Ст2пс
Механические свойства по стандарту ГОСТ 380-2005
|
Предел текучести, σ0,2, МПа |
Временное сопротивление разрыву, σв, МПа |
Относительное удлинение при разрыве, δ5 |
| 195 – 225 | 335 – 430 |
29 – 32 |
Механические свойства по стандарту ГОСТ 535-2005
|
Состояние поставки |
Категория |
Толщина проката, мм |
Предел текучести, σ0,2, МПа | Временное сопротивление разрыву, σв, МПа | Относительное удлинение при разрыве, δ5, % | Изгиб до параллельности сторон |
| Прокат |
1 |
< 20 |
> 225 |
335 – 430 |
> 32 |
d = a |
| Прокат |
1 |
20 – 40 |
> 215 |
335 – 430 |
> 31 |
d = 2a |
| Прокат |
1 |
40 – 100 |
> 205 |
335 – 430 |
> 29 |
d = 2a |
| Прокат |
1 |
> 100 |
> 195 |
335 – 430 |
> 29 |
d = 2a |
| Прокат |
По запросу |
– |
– |
> 335 |
– |
– |
|
Фасонный прокат |
||||||
|
Прокат |
– |
< 20 |
– |
– |
> 31 |
– |
|
Прокат |
– |
20 – 40 |
> 205 |
– |
> 30 |
– |
|
Прокат |
– |
40 – 100 |
> 195 |
– |
> 28 |
– |
|
Прокат |
– |
> 100 |
> 185 |
– |
> 28 |
– |
Механические свойства по стандарту ГОСТ 30136-95
|
Сортамент |
Охлаждение |
Временное сопротивление разрыву, σв, МПа |
Изгиб до параллельности сторон |
Относительное сужение, ψ, % |
|
Катанка |
Одностадийное |
< 420 |
d = a |
> 60 |
|
Катанка |
Двухстадийное |
< 470 |
d = a |
> 60 |
|
Катанка |
На воздухе |
< 420 |
d = a |
> 60 |
×
Отмена Удалить
×
Выбрать тариф
×
Подтверждение удаления
Отмена Удалить
×
Выбор региона будет сброшен
Отмена
×
×
Оставить заявку
×
| Название | |||
Отмена
×
К сожалению, данная функция доступна только на платном тарифе
Выбрать тариф
Сталь конструкционная углеродистая обыкновенного качества Ст2пс – характеристики, свойства, аналоги
На данной страничке приведены технические, механические и остальные свойства, а также характеристики стали марки Ст2пс.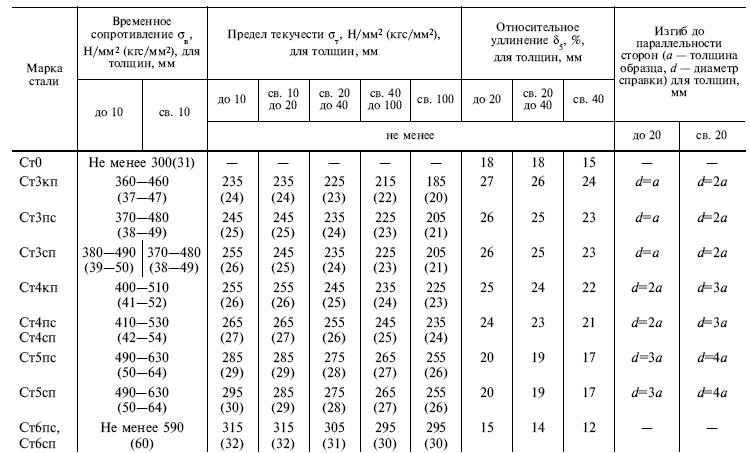
Марка: Ст2пс
Классификация материала: Сталь конструкционная углеродистая обыкновенного качества
Применение: неответственные детали, требующие повышенной пластичности или глубокой вытяжки- маланагруженные элементы сварных конструкций, работающие при постоянных нагрузках и при положительных температурах
Химический состав материала Ст2пс в процентном соотношении
| C | Si | Mn | Ni | S | P | Cr | N | Cu | As |
| 0.09 – 0.15 | 0.05 – 0.15 | 0.25 – 0.5 | до 0.3 | до 0.05 | до 0.04 | до 0.3 | до 0.008 | до 0.3 | до 0.08 |
Механические свойства Ст2пс при температуре 20
oС| Сортамент | Размер | Напр. | sв | sT | d5 | y | KCU | Термообр. |
| – | мм | – | МПа | МПа | % | % | кДж / м2 | – |
| Трубы, ГОСТ 8696-74 | 334 | 225 | 24 | |||||
| Трубы, ГОСТ 10705-80 | 333 | 206 | 24 | |||||
| Прокат, ГОСТ 535-2005 | 335-430 | 195-225 | 29-32 | |||||
| Лист толстый, ГОСТ 14637-89 | 320-410 | 185-215 | 30-33 | |||||
| Катанка, ГОСТ 30136-95 | 420-470 | 60 |
Технологические свойства Ст2пс
| Свариваемость: | без ограничений. |
| Флокеночувствительность: | не чувствительна. |
| Склонность к отпускной хрупкости: | не склонна. |
Расшифровка обозначений, сокращений, параметров
| Механические свойства : | |
| sв | – Предел кратковременной прочности , [МПа] |
| sT | – Предел пропорциональности (предел текучести для остаточной деформации), [МПа] |
| d5 | – Относительное удлинение при разрыве , [ % ] |
| y | – Относительное сужение , [ % ] |
| KCU | – Ударная вязкость , [ кДж / м2] |
| HB | – Твердость по Бринеллю , [МПа] |
Физические свойства : | |
| T | – Температура, при которой получены данные свойства , [Град] |
| E | – Модуль упругости первого рода , [МПа] |
| a | – Коэффициент температурного (линейного) расширения (диапазон 20o– T ) , [1/Град] |
| l | – Коэффициент теплопроводности (теплоемкость материала) , [Вт/(м·град)] |
| r | – Плотность материала , [кг/м3] |
| C | – Удельная теплоемкость материала (диапазон 20o– T ), [Дж/(кг·град)] |
| R | – Удельное электросопротивление, [Ом·м] |
Свариваемость : | |
| без ограничений | – сварка производится без подогрева и без последующей термообработки |
| ограниченно свариваемая | – сварка возможна при подогреве до 100-120 град. и последующей термообработке и последующей термообработке |
| трудносвариваемая | – для получения качественных сварных соединений требуются дополнительные операции: подогрев до 200-300 град. при сварке, термообработка после сварки – отжиг |
Другие марки из этой категории:
- Марка ВСт2кп
- Марка ВСт2пс
- Марка ВСт2сп
- Марка ВСт3Гпс
- Марка ВСт3кп
- Марка ВСт3пс
- Марка ВСт3сп
- Марка ВСт4кп
- Марка ВСт4пс
- Марка ВСт5пс
- Марка ВСт5сп
- Марка ВСт6пс
- Марка ВСт6сп
- Марка Ст0
- Марка Ст1
- Марка Ст1кп
- Марка Ст1пс
- Марка Ст1сп
- Марка Ст2кп
- Марка Ст2пс
- Марка Ст2сп
- Марка Ст3Гпс
- Марка Ст3Гсп
- Марка Ст3кп
- Марка Ст3пс
- Марка Ст3сп
- Марка Ст4кп
- Марка Ст4пс
- Марка Ст4сп
- Марка Ст5Гпс
- Марка Ст5пс
- Марка Ст5сп
- Марка Ст6пс
- Марка Ст6сп
Обращаем ваше внимание на то, что данная информация о марке Ст2пс, приведена в ознакомительных целях. Параметры, свойства и состав
реального материала марки Ст2пс могут отличаться от значений, приведённых на данной странице. Более подробную информацию о марке Ст2пс можно уточнить на
информационном ресурсе Марочник стали и сплавов. Информацию о наличии, сроках поставки и стоимости материалов Вы можете уточнить у наших менеджеров.
При обнаружении неточностей в описании материалов или найденных ошибках просим сообщать администраторам сайта, через форму обратной связи. Заранее спасибо за сотрудничество!
Параметры, свойства и состав
реального материала марки Ст2пс могут отличаться от значений, приведённых на данной странице. Более подробную информацию о марке Ст2пс можно уточнить на
информационном ресурсе Марочник стали и сплавов. Информацию о наличии, сроках поставки и стоимости материалов Вы можете уточнить у наших менеджеров.
При обнаружении неточностей в описании материалов или найденных ошибках просим сообщать администраторам сайта, через форму обратной связи. Заранее спасибо за сотрудничество!
Gardener’s Blue Ribbon ST2-2 Прочный кол, длина 2 фута, диаметр 5/16 дюйма, сталь, 20 шт. в упаковке #VORG4384913, ST2
Ремонт дома с 1928 годаМои товары
- Дом
 org/ListItem”>
Газон и сад
org/ListItem”>
Газон и сад- Товары для ландшафта
- Кашпо, аксессуары для растений
- Опоры растений
Артикул №: VORG4384913 – UPC: 048307700025 – МОДЕЛЬ №: ST2
Вещь #: ВОРГ4384913
Распродажа: $9,92
Розница: $23,00
Было: $21,60
Вы экономите: $13,08 (56,9%)
Количество:
MAT – MIDWEST AIR TECHNOLOGY
Об этом изделии
Gardener’s Blue Ribbon ST2-2 Прочный кол, длина 2 фута, диаметр 5/16 дюйма, сталь, 20 шт. в упаковке
в упаковке
- Стальной сердечник для долговечной работы
- Прочное толстое пластиковое покрытие
- Устойчивость к атмосферным воздействиям, химическим веществам и коррозии
- Поверхность с надежным захватом
Нержавеющий и с пластиковым покрытием для водонепроницаемой защиты, стальной сердечник для прочности. Шпоры предотвращают скольжение.
Длина. футов x диам.=2 x 5/16
СТАЛЬНОЙ СЕРДЕЧНИК
Единица измерения: шт.
- Диаметр: 5/16
- Длина: 2 дюйма
- Материал: сталь
Синяя лента Gardener’s ST5 Прочный кол, длина 5 футов, диаметр 7/16 дюйма, сталь, 20 шт. в упаковке
Распродажа: $24,52
Колья для растений Gardener’s Blue Ribbon BB4, 4 фута L, бамбук, натуральный бамбук
Цена: $5,26
Галстук для растений Gardener’s Blue Ribbon T001B, 100 футов L, пластик
Цена: 4,47 доллара США.

Gardener’s Blue Ribbon WW5-1 Садовый кол, 5 футов L, 0,66 дюйма W, дерево, натуральный, 24 упаковки
Распродажа: $63,14
Gardener’s Blue Ribbon ST3-3 Прочный кол, длина 3 фута, диаметр 5/16 дюйма, сталь, 20 шт. в упаковке
Распродажа: $13,22
Синяя лента Gardener’s ST5 Прочный кол, длина 5 футов, диаметр 7/16 дюйма, сталь, 20 шт.
 в упаковке
в упаковкеРаспродажа: $24,52
404 – СТРАНИЦА НЕ НАЙДЕНА
Почему я вижу эту страницу?
404 означает, что файл не найден. Если вы уже загрузили файл, имя может быть написано с ошибкой или файл находится в другой папке.
Другие возможные причиныВы можете получить ошибку 404 для изображений, поскольку у вас включена защита от горячих ссылок, а домен отсутствует в списке авторизованных доменов.
Если вы перейдете по временному URL-адресу (http://ip/~username/) и получите эту ошибку, возможно, проблема связана с набором правил, хранящимся в файле .htaccess. Вы можете попробовать переименовать этот файл в .htaccess-backup и обновить сайт, чтобы посмотреть, решит ли это проблему.
Также возможно, что вы непреднамеренно удалили корневую папку документа или вам может потребоваться повторное создание вашей учетной записи. В любом случае, пожалуйста, немедленно свяжитесь с вашим веб-хостингом.
В любом случае, пожалуйста, немедленно свяжитесь с вашим веб-хостингом.
Вы используете WordPress? См. Раздел об ошибках 404 после перехода по ссылке в WordPress.
Как найти правильное написание и папку
Отсутствующие или поврежденные файлыКогда вы получаете ошибку 404, обязательно проверьте URL-адрес, который вы пытаетесь использовать в своем браузере. Это сообщает серверу, какой ресурс он должен использовать попытка запроса.
http://example.com/example/Example/help.html
В этом примере файл должен находиться в папке public_html/example/Example/
Обратите внимание, что в этом примере важен CaSe . На платформах с учетом регистра e xample и E xample не совпадают.
Для дополнительных доменов файл должен находиться в папке public_html/addondomain.com/example/Example/, а имена чувствительны к регистру.
Неработающее изображение Если на вашем сайте отсутствует изображение, вы можете увидеть на своей странице поле с красным размером X , где отсутствует изображение. Щелкните правой кнопкой мыши на X и выберите «Свойства». Свойства сообщат вам путь и имя файла, который не может быть найден.
Щелкните правой кнопкой мыши на X и выберите «Свойства». Свойства сообщат вам путь и имя файла, который не может быть найден.
Это зависит от браузера. Если вы не видите на своей странице поле с красным X , попробуйте щелкнуть правой кнопкой мыши страницу, затем выберите «Просмотр информации о странице» и перейдите на вкладку «Мультимедиа».
http://example.com/cgi-sys/images/banner.PNG
В этом примере файл изображения должен находиться в папке public_html/cgi-sys/images/
Обратите внимание, что в этом пример. На платформах, которые обеспечивают чувствительность к регистру PNG и png — это не одно и то же место.
Ошибки 404 после перехода по ссылкам WordPress
При работе с WordPress ошибки 404 Page Not Found часто могут возникать при активации новой темы или изменении правил перезаписи в файле .htaccess.
Когда вы сталкиваетесь с ошибкой 404 в WordPress, у вас есть два варианта ее исправления.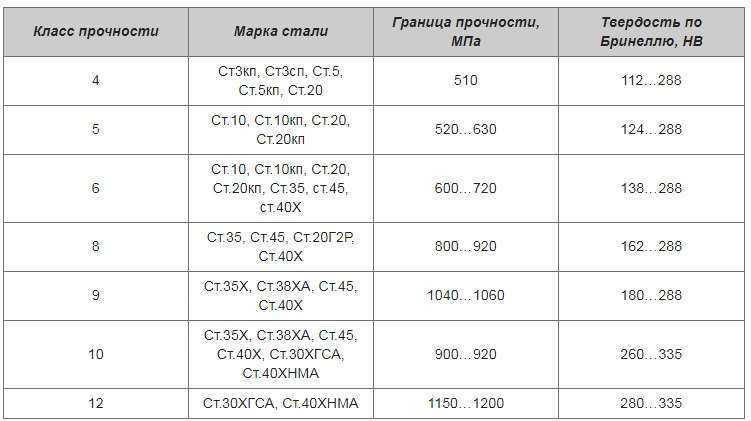
- Войдите в WordPress.
- В меню навигации слева в WordPress нажмите Настройки > Постоянные ссылки (Обратите внимание на текущую настройку. Если вы используете настраиваемую структуру, скопируйте или сохраните ее где-нибудь.)
- Выберите По умолчанию .
- Нажмите Сохранить настройки .
- Верните настройки к предыдущей конфигурации (до того, как вы выбрали «По умолчанию»). Верните пользовательскую структуру, если она у вас была.
- Нажмите Сохранить настройки .
Во многих случаях это сбросит постоянные ссылки и устранит проблему. Если это не сработает, вам может потребоваться отредактировать файл .htaccess напрямую.
Вариант 2. Измените файл .htaccessДобавьте следующий фрагмент кода в начало файла .htaccess:
# НАЧАЛО WordPress
 c>
c>
RewriteEngine On
RewriteBase / 9index.php$ – [L]
RewriteCond %{REQUEST_FILENAME} !-f
RewriteCond %{REQUEST_FILENAME} !-d
RewriteRule . /index.php [L]
# Конец WordPress
Если ваш блог показывает неправильное доменное имя в ссылках, перенаправляет на другой сайт или отсутствуют изображения и стиль, все это обычно связано с одной и той же проблемой: в вашем блоге WordPress настроено неправильное доменное имя.
Как изменить файл .htaccess
Файл .htaccess содержит директивы (инструкции), которые сообщают серверу, как вести себя в определенных сценариях, и напрямую влияют на работу вашего веб-сайта.
Перенаправление и перезапись URL-адресов — это две очень распространенные директивы, которые можно найти в файле .htaccess, и многие скрипты, такие как WordPress, Drupal, Joomla и Magento, добавляют директивы в .htaccess, чтобы эти скрипты могли работать.
Возможно, вам потребуется отредактировать файл . htaccess в какой-то момент по разным причинам. В этом разделе рассматривается, как редактировать файл в cPanel, но не то, что может потребоваться изменить. статьи и ресурсы для этой информации.)
htaccess в какой-то момент по разным причинам. В этом разделе рассматривается, как редактировать файл в cPanel, но не то, что может потребоваться изменить. статьи и ресурсы для этой информации.)
- Отредактируйте файл на своем компьютере и загрузите его на сервер через FTP
- Использовать режим редактирования программы FTP
- Использовать SSH и текстовый редактор
- Используйте файловый менеджер в cPanel
Самый простой способ отредактировать файл .htaccess для большинства людей — через диспетчер файлов в cPanel.
Как редактировать файлы .htaccess в файловом менеджере cPanelПрежде чем что-либо делать, рекомендуется сделать резервную копию вашего веб-сайта, чтобы вы могли вернуться к предыдущей версии, если что-то пойдет не так.
Откройте файловый менеджер- Войдите в cPanel.
- В разделе «Файлы» щелкните значок File Manager .

- Установите флажок для Корень документа для и выберите доменное имя, к которому вы хотите получить доступ, в раскрывающемся меню.
- Убедитесь, что установлен флажок Показать скрытые файлы (точечные файлы) “.
- Нажмите Перейти . Файловый менеджер откроется в новой вкладке или окне.
- Найдите файл .htaccess в списке файлов. Возможно, вам придется прокрутить, чтобы найти его.
- Щелкните правой кнопкой мыши файл .htaccess и выберите Редактировать код в меню. Кроме того, вы можете щелкнуть значок файла .htaccess, а затем Редактор кода значок в верхней части страницы.
- Может появиться диалоговое окно с вопросом о кодировании. Просто нажмите Изменить , чтобы продолжить. Редактор откроется в новом окне.
- При необходимости отредактируйте файл.

- Нажмите Сохранить изменения в правом верхнем углу, когда закончите. Изменения будут сохранены.
- Протестируйте свой веб-сайт, чтобы убедиться, что ваши изменения были успешно сохранены. Если нет, исправьте ошибку или вернитесь к предыдущей версии, пока ваш сайт снова не заработает.
- После завершения нажмите Закрыть , чтобы закрыть окно диспетчера файлов.
Нарушенная активация IL-33/ST2 в децидуализирующихся стромальных клетках продлевает восприимчивость матки у женщин с невынашиванием беременности
1. Ван Х, Дей С.К. (2006)Дорожная карта имплантации эмбрионов: подсказки на моделях мышей. Обзоры природы Генетика 7: 185–199. [PubMed] [Google Scholar]
2. Bazer FW, Spencer TE, Johnson GA, Burghardt RC, Wu G (2009) Сравнительные аспекты имплантации. Воспроизведение 138: 195–209. [PubMed] [Google Scholar]
3.
Brosens JJ, Gellersen B (2006)Смерть или выживание – прогестерон-зависимые решения судьбы клеток в строме эндометрия человека. Журнал молекулярной эндокринологии
36: 389–398. [PubMed] [Google Scholar]
Журнал молекулярной эндокринологии
36: 389–398. [PubMed] [Google Scholar]
4. Геллерсен Б., Бросенс И.А., Бросенс Дж.Дж. (2007)Децидуализация человеческого эндометрия: механизмы, функции и клинические перспективы. Семинары по репродуктивной медицине 25: 445–453. [PubMed] [Google Scholar]
5. Геллерсен Б., Бросенс Дж. (2003)Взаимодействие циклического АМФ и рецептора прогестерона в эндометрии человека: децидуализирующее дело. Журнал эндокринологии 178: 357–372. [PubMed] [Академия Google]
6. Achache H, Revel A (2006)Маркеры рецептивности эндометрия, путь к успешной имплантации эмбриона. Обновление репродукции человека 12: 731–746. [PubMed] [Google Scholar]
7. Aghajanova L, Hamilton AE, Giudice LC (2008) Восприимчивость матки к имплантации человеческого эмбриона: гистология, биомаркеры и транскриптомика. Семинары по биологии клетки и развития 19: 204–211. [Бесплатная статья PMC] [PubMed] [Google Scholar]
8.
Diedrich K, Fauser BC, Devroey P, Griesinger G (2007) Роль эндометрия и эмбриона в имплантации человека. Обновление репродукции человека
13: 365–377. [PubMed] [Академия Google]
Обновление репродукции человека
13: 365–377. [PubMed] [Академия Google]
9. Дей С.К., Лим Х., Дас С.К., Риз Дж., Париа Б.К. и др. (2004) Молекулярные сигналы имплантации. Эндокринные обзоры 25: 341–373. [PubMed] [Google Scholar]
10. Harper MJ (1992) Окно имплантации. Клиническое акушерство и гинекология Байьера 6: 351–371. [PubMed] [Google Scholar]
11. Эверс Дж. Л. (2002) Женское бесплодие. Ланцет 360: 151–159. [PubMed] [Google Scholar]
12. Текленбург Г., Салкер М., Хейнен С., Маклон Н.С., Бросенс Дж.Дж. (2010)Молекулярная основа привычного невынашивания беременности: нарушение естественного отбора эмбрионов. Молекулярная репродукция человека 16: 886–895. [PubMed] [Google Scholar]
13.
Салкер М., Текленбург Г., Молохия М., Лавери С., Трю Г. и др. (2010) Естественный отбор эмбрионов человека: нарушение децидуализации эндометрия отключает взаимодействие эмбриона и матери и вызывает повторяющееся невынашивание беременности. PloS один
5: e10287. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
14. Wilcox AJ, Baird DD, Weinberg CR (1999)Время имплантации зачатия и потеря беременности. Медицинский журнал Новой Англии 340: 1796–1799. [PubMed] [Академия Google]
15. Cloke B, Huhtinen K, Fusi L, Kajihara T, Yliheikkila M, et al. (2008) Рецепторы андрогенов и прогестерона регулируют различные генные сети и клеточные функции при децидуализации эндометрия. Эндокринология 149: 4462–4474. [Бесплатная статья PMC] [PubMed] [Google Scholar]
16. Джонс М.С., Фуси Л., Хайэм Дж.Х., Абдель-Хафиз Х., Хорвиц К.Б. и др. (2006) Регуляция пути SUMO повышает чувствительность дифференцирующихся стромальных клеток эндометрия человека к прогестерону. Труды Национальной академии наук Соединенных Штатов Америки 103: 16272–16277. [Бесплатная статья PMC] [PubMed] [Google Scholar]
17.
Аль-Саббах М., Фуси Л., Хайэм Дж., Ли И., Лей К. и др. (2011) Активные формы кислорода, полученные из NADPH-оксидазы, опосредуют децидуализацию стромальных клеток эндометрия человека в ответ на передачу сигналов циклического AMP. Эндокринология
152: 730–740. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Эндокринология
152: 730–740. [Бесплатная статья PMC] [PubMed] [Google Scholar]
18. Ван С., Танака Т., Накамура Х., Умесаки Н., Хираи К. и др. (2003)Гранулированные клетки метриальной железы в мышиной матке: локализация, кинетика и функциональная роль в ангиогенезе во время беременности. Микроскопические исследования и техника 60: 420–429. [PubMed] [Google Scholar]
19. Чжан Дж., Чен З., Смит Г.Н., Крой Б.А. (2011)Трансформация сосудов, запускаемая естественными клетками-киллерами: материнская забота до рождения? Клеточная и молекулярная иммунология 8: 1–11. [Бесплатная статья PMC] [PubMed] [Google Scholar]
20. Ласкарин Г., Стрбо Н., Богович Крнчич Т., Юретич К., Леди Батай Н. и др. (2005)Физиологическая роль IL-15 и IL-18 в контакте матери и плода. Химическая иммунология и аллергия 89: 10–25. [PubMed] [Академия Google]
21.
Нэнси П., Тальяни Э., Тай К.С., Асп П., Леви Д.Э. и соавт. (2012)Сайленсинг хемокинового гена в децидуальных стромальных клетках ограничивает доступ Т-клеток к материнско-плодовому интерфейсу. Наука
336: 1317–1321. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Наука
336: 1317–1321. [Бесплатная статья PMC] [PubMed] [Google Scholar]
22. Guzeloglu-Kayisli O, Kayisli UA, Taylor HS (2009)Роль факторов роста и цитокинов во время имплантации: эндокринные и паракринные взаимодействия. Семинары по репродуктивной медицине 27: 62–79. [Бесплатная статья PMC] [PubMed] [Google Scholar]
23. Wegmann TG, Lin H, Guilbert L, Mosmann TR (1993)Двунаправленные взаимодействия цитокинов в отношениях матери и плода: является ли успешная беременность феноменом Th3? Иммунология сегодня 14: 353–356. [PubMed] [Google Scholar]
24. Chaouat G, Tranchot Diallo J, Volumenie JL, Menu E, Gras G и др. (1997) Иммунная супрессия и баланс Th2/Th3 во время беременности: (очень) личная дань уважения Тому Вегманну. Американский журнал репродуктивной иммунологии 37: 427–434. [PubMed] [Академия Google]
25.
Wilczynski JR (2005)Баланс цитокинов Th2/Th3 – инь и ян репродуктивной иммунологии. Европейский журнал акушерства, гинекологии и репродуктивной биологии
122: 136–143. [PubMed] [Google Scholar]
[PubMed] [Google Scholar]
26. Kakkar R, Lee RT (2008)Путь IL-33/ST2: терапевтическая цель и новый биомаркер. Обзоры природы Открытие лекарств 7: 827–840. [Бесплатная статья PMC] [PubMed] [Google Scholar]
27. Миллер А.М., Лью Ф.Ю. (2011)Путь IL-33/ST2 – новая терапевтическая мишень при сердечно-сосудистых заболеваниях. Фармакология и терапия 131: 179–186. [PubMed] [Google Scholar]
28. Палмер Г., Габай С. (2011) Биология интерлейкина-33 с потенциальным пониманием болезней человека. Отзывы о природе Ревматология 7: 321–329. [PubMed] [Google Scholar]
29. Haraldsen G, Balogh J, Pollheimer J, Sponheim J, Kuchler AM (2009) Интерлейкин-33 – цитокин двойной функции или новый алармин? Тенденции в иммунологии 30: 227–233. [PubMed] [Google Scholar]
30.
Карьер В., Руссель Л., Ортега Н., Лакорре Д.А., Америх Л. и др. (2007) IL-33, IL-1-подобный цитокиновый лиганд для рецептора ST2, представляет собой хроматин-ассоциированный ядерный фактор in vivo. Труды Национальной академии наук Соединенных Штатов Америки
104: 282–287. [Бесплатная статья PMC] [PubMed] [Google Scholar]
[Бесплатная статья PMC] [PubMed] [Google Scholar]
31. Палмер Г., Липски Б.П., Смитгалл М.Д., Майнингер Д., Сиу С. и соавт. (2008) Дополнительный белок рецептора IL-1 (AcP) необходим для передачи сигналов IL-33, а растворимый AcP усиливает способность растворимого ST2 ингибировать IL-33. Цитокин 42: 358–364. [PubMed] [Google Scholar]
32. Миллер А.М. (2011)Роль IL-33 в воспалении и заболевании. Журнал воспаления 8: 22. [Бесплатная статья PMC] [PubMed] [Google Scholar]
33. Brosens JJ, Hayashi N, White JO (1999) Рецептор прогестерона регулирует децидуальную экспрессию пролактина в дифференцирующихся стромальных клетках эндометрия человека. Эндокринология 140: 4809–4820. [PubMed] [Google Scholar]
34. Кононен Дж., Бубендорф Л., Каллиониеми А., Барлунд М., Шрамл П. и соавт. (1998)Тканевые микрочипы для высокопроизводительного молекулярного профилирования образцов опухолей. Природная медицина 4: 844–847. [PubMed] [Google Scholar]
35.
Такано М., Лу З. , Гото Т., Фуси Л., Хайэм Дж. и др. (2007) Транскрипционная перекрестная связь между фактором транскрипции forkhead box O1A и рецептором прогестерона координирует регуляцию клеточного цикла и дифференцировку в стромальных клетках эндометрия человека. Молекулярная эндокринология
21: 2334–2349. [PubMed] [Google Scholar]
, Гото Т., Фуси Л., Хайэм Дж. и др. (2007) Транскрипционная перекрестная связь между фактором транскрипции forkhead box O1A и рецептором прогестерона координирует регуляцию клеточного цикла и дифференцировку в стромальных клетках эндометрия человека. Молекулярная эндокринология
21: 2334–2349. [PubMed] [Google Scholar]
36. Шервин Дж. Р., Шарки А. М., Камео П., Маврогианис П. М., Каталано Р. Д. и др. (2007)Идентификация новых генов, регулируемых хорионическим гонадотропином, в эндометрии павианов во время окна имплантации. Эндокринология 148: 618–626. [PubMed] [Google Scholar]
37. Salker MS, Christian M, Steel JH, Nautiyal J, Lavery S, et al.. (2011) Дерегуляция сывороточной и глюкокортикоид-индуцируемой киназы SGK1 в эндометрии вызывает репродуктивную недостаточность . Природная медицина. [ПубМед]
38.
Сингх М., Чаудхри П., Асселин Э. (2011)Связывание восприимчивости эндометрия и имплантации: сеть гормонов, цитокинов и факторов роста. Журнал эндокринологии
210: 5–14. [PubMed] [Google Scholar]
[PubMed] [Google Scholar]
39. Koot YE, Teklenburg G, Salker MS, Brosens JJ, Macklon NS (2012)Молекулярные аспекты неудачи имплантации. Биохимика и биофизика акта. [PubMed]
40. Matsumoto H, Zhao X, Das SK, Hogan BL, Dey SK (2002)Индийский еж как чувствительный к прогестерону фактор, опосредующий эпителиально-мезенхимальные взаимодействия в матке мыши. Биология развития 245: 280–290. [PubMed] [Google Scholar]
41. Такамото Н., Чжао Б., Цай С.Ю., ДеМайо Ф.Дж. (2002)Идентификация индийского ежа как гена, чувствительного к прогестерону, в мышиной матке. Молекулярная эндокринология 16: 2338–2348. [PubMed] [Google Scholar]
42. Li Q, Kannan A, DeMayo FJ, Lydon JP, Cooke PS, et al. (2011) Антипролиферативное действие прогестерона на эпителий матки опосредовано Hand2. Наука 331: 912–916. [Бесплатная статья PMC] [PubMed] [Google Scholar]
43.
Nallasamy S, Li Q, Bagchi MK, Bagchi IC (2012)Гены гомеобокса Msx критически регулируют имплантацию эмбриона, контролируя паракринную передачу сигналов между стромой матки и эпителием. Генетика PLoS
8: е1002500. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Генетика PLoS
8: е1002500. [Бесплатная статья PMC] [PubMed] [Google Scholar]
44. Лью Ф.Ю., Питман Н.И., Макиннес И.Б. (2010)Функции IL-33, связанные с заболеванием: новый ребенок в семье IL-1. Обзоры природы Иммунология 10: 103–110. [PubMed] [Google Scholar]
45. Чжао В., Ху З. (2010) Загадочная обработка и секреция интерлейкина-33. Клеточная и молекулярная иммунология 7: 260–262. [Бесплатная статья PMC] [PubMed] [Google Scholar]
46. Kakkar R, Hei H, Dobner S, Lee RT (2012) Интерлейкин 33 как механически чувствительный цитокин, секретируемый живыми клетками. Журнал биологической химии 287: 6941–6948. [Бесплатная статья PMC] [PubMed] [Google Scholar]
47. Хосино К., Кашивамура С., Курибаяши К., Кодама Т., Цудзимура Т. и др. (1999) Отсутствие T1/ST2, связанных с рецептором интерлейкина 1, не влияет на развитие Т-хелперных клеток 2 типа и их эффекторную функцию. Журнал экспериментальной медицины 190: 1541–1548. [Бесплатная статья PMC] [PubMed] [Google Scholar]
48. Гранн И., Сауткомб Дж. Х., Снайдер Дж. В., Таннетта Д. С., Чайлд Т. и др. (2011) ST2 и IL-33 при беременности и преэклампсии. PloS один
6: е24463. [Бесплатная статья PMC] [PubMed] [Google Scholar]
Гранн И., Сауткомб Дж. Х., Снайдер Дж. В., Таннетта Д. С., Чайлд Т. и др. (2011) ST2 и IL-33 при беременности и преэклампсии. PloS один
6: е24463. [Бесплатная статья PMC] [PubMed] [Google Scholar]
49. Kaitu’u-Lino TJ, Tuohey L, Tong S (2012)Интерлейкин-33 материнской сыворотки и растворимый ST2 на ранних сроках беременности и их связь с выкидышем. Журнал репродуктивной иммунологии 95: 46–49. [PubMed] [Google Scholar]
50. Quenby S, Vince G, Farquharson R, Aplin J (2002) Повторяющийся выкидыш: дефект в природном контроле качества? Репродукция человека 17: 1959–1963. [PubMed] [Google Scholar]
51. Рай Р., Риган Л. (2006) Повторяющийся выкидыш. Ланцет 368: 601–611. [PubMed] [Академия Google]
52. Grimaldi G, Christian M, Quenby S, Brosens JJ (2012)Экспрессия эпигенетических эффекторов в децидуализирующихся стромальных клетках эндометрия человека. Молекулярная репродукция человека. [PubMed]
53.
Gargett CE, Ye L (2012)Реконструкция эндометрия из стволовых клеток. Фертильность и бесплодие
98: 11–20. [PubMed] [Google Scholar]
Фертильность и бесплодие
98: 11–20. [PubMed] [Google Scholar]
54. Чапюи Дж., Хот Д., Хансманнель Ф., Кердраон О., Феррейра С. и др. (2009) Транскриптомные и генетические исследования идентифицируют IL-33 как ген-кандидат для болезни Альцгеймера. Молекулярная психиатрия 14: 1004–1016. [Бесплатная статья PMC] [PubMed] [Google Scholar]
55. Humphreys NE, Xu D, Hepworth MR, Liew FY, Grencis RK (2008) IL-33, мощный индуктор адаптивного иммунитета к кишечным нематодам. Журнал иммунологии 180: 2443–2449. [PubMed] [Google Scholar]
56. Kunes P, Holubcova Z, Kolackova M, Krejsek J (2010)Интерлейкин-33, новый член семейства цитокинов IL-1/IL-18, в кардиологии и кардиохирургии. Торакальный и сердечно-сосудистый хирург 58: 443–449. [PubMed] [Google Scholar]
57.
Миллер А.М., Асквит Д.Л., Хьюбер А.Дж., Андерсон Л.А., Холмс В.М. и др. (2010) Интерлейкин-33 оказывает защитное действие при воспалении жировой ткани при ожирении у мышей. Исследование тиража
107: 650–658.