Плотность р18: характеристики и расшифовка, применение и свойства стали
alexxlab | 29.05.1984 | 0 | Разное
Р18
Главная/Характеристики Марок Стали, Металлопроката/Р18
Характеристика материала. Сталь Р18.
| Марка | Сталь Р18 |
| Заменитель | СТАЛЬ Р12 |
| Классификация | СТАЛЬ ИНСТРУМЕНТАЛЬНАЯ БЫСТРОРЕЖУЩАЯ (БЫСТРОРЕЗ НОРМАЛЬНОЙ ПРОИЗВОДИТЕЛЬНОСТИ) |
| Прочие обозначения | |
| Иностранные аналоги | AISI T1, DIN HS18-0-1 |
| Применение |  |
| Вид поставки | |
| Сортовой прокат, в том числе фасонный: | ГОСТ 2590-88, ГОСТ 2591-88 |
| Калиброванный пруток | ГОСТ 19265-73, ГОСТ 7417-75 |
| Шлифованный пруток и серебрянка | ГОСТ 19265-73, ГОСТ 14955-77 |
| Лист толстый | ТУ 14-1-1408-75 |
| Лист тонкий | ТУ 14-1-1408-75, ТУ 14-1-1706-76 |
| Лента | ГОСТ 4986-79 |
| Полоса | ГОСТ 19265-73, ГОСТ 4405-75 |
| Проволока | ТУ 14-1-1096-74 |
| Поковки и кованые заготовки | ГОСТ 19265-73, ГОСТ 1133-71, ГОСТ 4405-75 |
Краткое описание и применение.
Быстрорежущая сталь Р18 относится к быстрорежущим сталям нормальной производительности. Эта сталь, содержащая 18% вольфрама, долгое время была наиболее распространенной. Инструменты, изготовленные из этой стали, после термической обработки имеют твердость HRC 62-65, красностойкость 600о С и достаточно высокую прочность. Сталь Р18 сравнительно хорошо шлифуется. Существенным недостатком этой стали является большая карбидная неоднородность, особенно значительная в прутках большого сечения.
При увеличении карбидной неоднородности прочность стали снижается и при работе наблюдается выкрашивание режущих кромок инструмента и снижение его стойкости.
Большое количество избыточной карбидной фазы делает сталь Р18 более мелкозернистой, менее чувствительной к перегреву при закалке, более износостойкой. Из стали Р18 могут изготовляться всевозможные инструменты, в том числе такие сложные как шеверы, долбяки, протяжки и др.
Сталь Р18 относится к быстрорежущим сталям нормальной теплостойкости. Быстрорежущие стали наиболее характерны для режущих инструментов. Они сочетают высокую теплостойкость (500-650°С) в зависимости от состава и обработки) с высокими твердостью (до HRC 68-70), износостойкостью при повышенных температурах и повышенным сопротивлением пластической деформации. Быстрорежущие стали позволяют повысить скорость резания в 2 – 4 раза по сравнению со скоростями, применяемыми при обработке инструментами из углеродистых и легированных инструментальных сталей.
Быстрорежущие стали наиболее характерны для режущих инструментов. Они сочетают высокую теплостойкость (500-650°С) в зависимости от состава и обработки) с высокими твердостью (до HRC 68-70), износостойкостью при повышенных температурах и повышенным сопротивлением пластической деформации. Быстрорежущие стали позволяют повысить скорость резания в 2 – 4 раза по сравнению со скоростями, применяемыми при обработке инструментами из углеродистых и легированных инструментальных сталей.
Высокие режущие свойства быстрорежущих сталей обеспечиваются легированием сильными карбидообразующими элементами (вольфрамом, молибденом, ванадием), элементами, повышающими температуру ( ‘ )-превращения (кобальтом, алюминием), и применением специальной термической обработки, заключающейся в закалке с высоких температур (1200 – 1300 °С) и отпуске, вызывающем дисперсионное твердение.

Для стали Р18 основным является карбид М6С (Fe3W3C).
Для получения высоких теплостойкости и твердости достаточно большая доля распадающегося карбида должна быть переведена при закалке в твердый раствор (аустенит, мартенсит), что насыщает его углеродом, вольфрамом, молибденом, ванадием, хромом.
Последующий отпуск при температурах 550-560 °C повышает твердость до максимальных значений вследствие выделения дисперсных, карбидов и распада остаточного аустенита.
В зависимости от химического состава, а следовательно, и уровня основных свойств быстрорежущие стали подразделяют на стали нормальной и повышенной теплостойкости (производительности). Если содержание ванадия не превышает 2%, их относят к быстрорежущим сталям нормальной теплостойкости (производительности). Это стали Р18, Р9 , Р6М5.
Быстрорежущие стали с более высоким содержанием ванадия, а также дополнительно легированные кобальтом относят к сталям повышенной теплостойкости (Р12Ф3 , Р6М5Ф3 , Р18Ф3К5 , Р9К5 , Р6М5К5 , Р9М4К8 и др.
 ).
).К группе быстрорежущих сталей повышенной производительности следует отнести и быстрорежущие дисперсионно-твердеющие сплавы с интерметаллидным упрочнением. Их высокая теплостойкость и режущие свойства обеспечиваются высокими температурами ( ‘ )-превращения и упрочнением вследствие выделения при отпуске интерметаллидов.
Для инструментов сложной формы, тонколезвийных, а также для инструментов, используемых при прерывистом точении, большее значение приобретают прочность и вязкость быстрорежущей стали.
Нагрев под закалку быстрорежущих сталей проводится при температурах и выдержках, обеспечивающих растворение специальных карбидов и легирование аустенита, для получения оптимальной теплостойкости. При закалке тонколезвийных инструментов небольших размеров (менее 3-5 мм) температура закалки снижается на 10-20 °С (балл зерна 11).
 Выдержку при окончательном нагреве выбирают из расчета 10-15 с на 1 мм диаметра (толщины) для инструмента диаметром 5-30 мм. Пусть толщина фрезы 3-5 мм, следовательно время выдержки при окончательном нагреве 40 c; температура – 1280 °С.
Выдержку при окончательном нагреве выбирают из расчета 10-15 с на 1 мм диаметра (толщины) для инструмента диаметром 5-30 мм. Пусть толщина фрезы 3-5 мм, следовательно время выдержки при окончательном нагреве 40 c; температура – 1280 °С.Время выдержки при подогреве обычно берут удвоенным по сравнению с выдержкой при окончательном нагреве.
Подогрев и окончательный нагрев под закалку быстрорежущих сталей, как правило, проводят в электродных соляных ваннах. Среда нагрева: при подогреве – смесь 78 % ВаС12 и 22 % NaCl (Тпл = 640 °С), при окончательном нагреве – в расплаве ВаС12 (Tпл = 962 °С). Для предохранения от окисления ванну раскисляют фтористым магнием.
Переохлажденный аустенит быстрорежущих сталей устойчив, вследствие чего они могут охлаждаться в любой среде – воздухе, масле, горячих средах при 500-560 °С. Будем охлаждать изделие в масле.
Отпуск быстрорежущих сталей выполняется при температурах 550-570 °С, 2-3 раза по 1 ч. Быстрорежущие стали с большой устойчивостью остаточного аустенита требуют трех- и даже четырехкратного отпуска.
 Будем выполнять трехкратный отпуск при температуре 550 °С.
Будем выполнять трехкратный отпуск при температуре 550 °С.При отпуске происходит выделение упрочняющих карбидов и распад остаточного аустенита. В результате быстрорежущая сталь получает высокую твердость, прочность и теплостойкость.
При закалке в аустените растворяется весь хром, 8% W, 1% V и 0,4-0,5% C. После закалки в структуре кроме мартенсита и первичных карбидов содержится 30-40% остаточного аустенита. Остаточный аустенит превращают в мартенсит при отпуске. Аустенит, обедняясь углеродом и легирующими элементами, становится менее устойчивым и при охлаждении ниже точки MН испытывает мартенситное превращение. Однократного отпуска недостаточно для превращения всего остаточного аустенита, поэтому применяют многократный отпуск.
Химический состав в % материала Р18
| C | Si | Mn | Ni | S | P | Cr | Mo | W | V | Co |
0. 73 – 0.83 73 – 0.83 | до 0.5 | до 0.5 | до 0.4 | до 0.03 | до 0.03 | 3.8 – 4.4 | до 1 | 17 – 18.5 | 1 – 1.4 | до 0.5 |
Механическиесвойствастали Р18
Механические свойства в зависимости от температуры отпуска
| tисп, 0С | σВ, Мпа | KCU, Дж/м2 | HRCэ |
Закалка 1280 °С, масло. Отпуск трехкратный по 1 ч. Отпуск трехкратный по 1 ч. | |||
| 400 | 1370 | 23 | 61 |
| 500 | 1470 | 19 | 63 |
| 550 | 2350 | 17 | 66 |
| 600 | 2210 | 65 | |
Твёрдость быстрореза Р18 после термообработки
| Твердость | Температура, °С | |||
| после отжига | после закалки с отпуском HRC3 (HRC), не менее | закалки | отпуска | |
| НВ, неболее | диаметр отпечатка, мм, не менее | |||
| 255 | 3,8 | 63(62) | 1270 | 560 |
Температура критических точек стали Р18
| Ar1=725 °С | Ar3(Arcm) =770 °С | Ac1=820 °С | Ac3=860 °С |
Физические свойства стали Р18
| Температура испытания,°С | 20 | 100 | 200 | 300 | 400 | 500 | 600 | 700 | 800 | 900 |
| Модуль нормальной упругости, Е, ГПа | 228 | 223 | 219 | 210 | 201 | 192 | 181 | |||
| Модуль упругости при сдвиге кручением G, ГПа | 83 | |||||||||
| Плотность, pn, кг/см3 | 8800 | |||||||||
| Коэффициент теплопроводности Вт/(м ·°С) | 26 | 27 | 28 | 29 | 28 | 27 | 27 | |||
Уд. электросопротивление (p, НОм · м) электросопротивление (p, НОм · м) | 419 | 472 | 544 | 627 | 718 | 815 | 922 | 1037 | 1152 | 1173 |
Красностойкость (теплостойкость) стали Р18
| Температура, °С | Время, ч | Твердость, HRCэ |
| 620 | 4 | 59 |
Технологические свойства материала Р18
| Температура ковки | Начала 1200, конца 900. Охлаждение в колодцах при 750-800 С. Охлаждение в колодцах при 750-800 С. |
| Свариваемость | при стыковой электросварке со сталью 45 и 40Х хорошая. |
| Шлифуемость | повышенная (ГОСТ 19265-73) |
| Обрабатываемость резанием | Kv тв.спл.= 0,6 и Kv б.ст = 0,3 резанием при HB 212-228 |
| Флокеночувствительность | не чувствительна |
Обозначения:
| Механические свойства | |
| sв | – Предел кратковременной прочности , [МПа] |
| sT | – Предел пропорциональности (предел текучести для остаточной деформации), [МПа] |
| d5 | – Относительное удлинение при разрыве , [ % ] |
| y | – Относительное сужение , [ % ] |
| KCU | – Ударная вязкость , [ кДж / м2] |
| HB | – Твердость по Бринеллю , [МПа] |
| Физические свойства : | |
| T | – Температура, при которой получены данные свойства , [Град] |
| E | – Модуль упругости первого рода , [МПа] |
| a | – Коэффициент температурного (линейного) расширения (диапазон 20o – T ) , [1/Град] |
| l | – Коэффициент теплопроводности (теплоемкость материала) , [Вт/(м·град)] |
| r | – Плотность материала , [кг/м3] |
| C | – Удельная теплоемкость материала (диапазон 20o – T ), [Дж/(кг·град)] |
| R | – Удельное электросопротивление, [Ом·м] |
| Свариваемость | |
| без ограничений | – сварка производится без подогрева и без последующей термообработки |
| ограниченно свариваемая | – сварка возможна при подогреве до 100-120 град.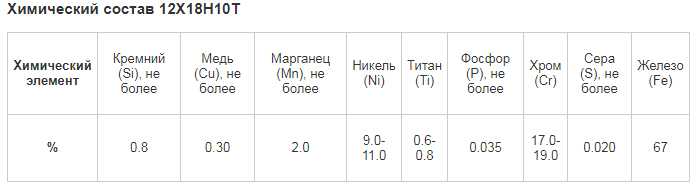 и последующей термообработке и последующей термообработке |
| трудносвариваемая | – для получения качественных сварных соединений требуются дополнительные операции: подогрев до 200-300 град. при сварке, термообработка после сварки – отжиг |
Купить сталь Р18. Быстрорежущая сталь со средним содержанием углерода 0,8% и легированная вольфрамом(18%), хромом(4%), ванадием(до 1.4%).
характеристики и расшифовка, применение и свойства стали
- Стали
- Стандарты
Всего сталей
| Страна | Стандарт | Описание | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Россия | ГОСТ 19265-73 | Прутки и полосы из быстрорежущей стали. Технические условия | ||||||||||
 js_elem_334344″>
js_elem_334344″>Характеристики стали Р18
| Классификация | Сталь инструментальная быстрорежущая |
| Применение | Резцы, сверла, фрезы, резьбовые фрезы, долбяки, развертки, зенкеры, метчики, протяжки для обработки конструкционных сталей с прочностью до 1000 МПа, от которых требуется сохранение режущих свойств при нагревании во время работы до 600 °С.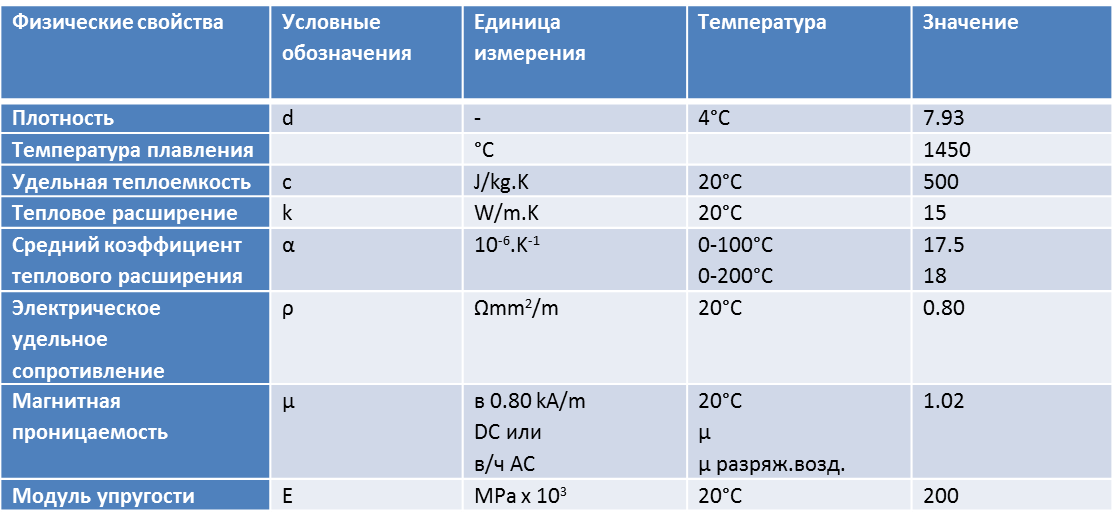 |
Сталь имеет пониженную склонность к перегреву при закалке, хорошую вязкость, хорошее сопротивление износу, повышенную шлифуемость
Механические свойства стали Р18
Механические свойства после отжига при повышенных температурах
| Температура испытания, °С | Предел текучести, σ0,2, МПа | Временное сопротивление разрыву, σв, МПа | Относительное удлинение при разрыве, δ5, % | Относительное сужение, ψ, % | Предел текучести при сжатии, σсж, МПа | Предел прочности при кручении, Jк, МПа | Ударная вязкость KCU при 20°С, Дж/см2 | Твердость, НВ |
| 200 | 450 | 830 | 13 | 22 | 1050 | 520 | - | 227 |
| 400 | 420 | 700 | 15 | 22 | 850 | 450 | - | 210 |
| 600 | 300 | 480 | 31 | 55 | 620 | 300 | - | 140 |
| 800 | 110 | 200 | 60 | 70 | 100 | 100 | - | 30 |
| 1000 | 90 | 100 | 42 | 55 | 50 | 50 | 100 | 24 |
| 1100 | - | - | - | - | - | - | 130 | - |
| 1200 | 30 | 30 | 12 | 25 | 40 | 40 | 45 | 4 |
Механические свойства в термообработанном состоянии при повышенных температурах
| Температура испытания, °С | Предел прочности при изгибе, σизг, МПа | Твердость по Виккерсу, HV | Твердость, HRC∂ |
| 200 | 3570 | 815 | 64 |
| 400 | 3730 | 755 | 62 |
| 500 | 3290 | 712 | 60 |
| 550 | 3060 | 661 | 58 |
| 600 | 2430 | 615 | 56 |
| 650 | 2180 | 504 | 51 |
Механические свойства в зависимости от температуры отпуска
| Температура отпуска, °С | Временное сопротивление разрыву, σв, МПа | Ударная вязкость KCU при 20°С, Дж/см2 | Твердость, HRC∂ |
Закалка при 1280 °С, масло. Отпуск трехкратный по 1 часу Отпуск трехкратный по 1 часу
|
|||
| 400 | 1370 | 23 | 61 |
| 500 | 1470 | 19 | 63 |
| 550 | 2350 | 17 | 66 |
| 600 | 2210 | - | 65 |
Свойства по стандарту ГОСТ 19265-73
|
Состояние |
Твердость, НВ (HRC) |
Диаметр отпечатка, мм |
|
После отжига |
< 225 |
> 3,8 |
|
После закалки с отпуском* |
> (62) |
– |
Физические свойства Р18
| Температура, °С |
Модуль упругости, 10– 5, МПа |
Коэффициент температурного расширения, 106, 1/°C | Коэффициент теплопроводности, Вт/м·°С | Удельная теплоемкость, Дж/кг·°С | Удельное электросопротивление, Ом·м |
| 20 |
2,28 |
– |
– |
– |
419 |
| 100 |
2,23 |
– |
26 |
– |
472 |
| 200 |
2,19 |
– |
27 |
– |
544 |
| 300 |
2,10 |
– |
28 |
– |
627 |
| 400 |
2,01 |
– |
29 |
– |
718 |
| 500 |
1,92 |
– |
28 |
– |
815 |
| 600 |
1,81 |
– |
27 |
– |
922 |
| 700 |
– |
– |
27 |
– |
1037 |
| 800 |
– |
– |
– |
– |
1152 |
| 900 |
– |
– |
– |
– |
1173 |
Свойства по стандарту ГОСТ 19265-73
Плотность, г/см3: 9,82*
*Типичное значение свойства для низкоуглеродистой и низколегированной
стали. Эта величина не предусмотрена стандартами, она носит
ориентировочный характер и не может быть использована с целью
проектирования.
Эта величина не предусмотрена стандартами, она носит
ориентировочный характер и не может быть использована с целью
проектирования.
×
Отмена Удалить
×
Выбрать тариф
×
Подтверждение удаления
Отмена Удалить
×
Выбор региона будет сброшен
Отмена
×
×
Оставить заявку
×
| Название | |||
Отмена
×
К сожалению, данная функция доступна только на платном тарифе
Выбрать тариф
размеры, вес 1 метра, основные характеристики R18 плотности по ГОСТу 6368 82
Содержание статьи
- Общие параметры
- Особенности изготовления
- Маркировка
- Схемы
- Сфера применения
- Расчетные значения размеров
- Хранение
- Транспортировка
Железнодорожное сообщение с успехом пользуется в самых разнообразных сферах промышленности, производства, поставок. И если стандартное полотно по типу Р65 зачастую необходимо для обычной транспортировки грузов между муниципальными образованиями, то конкретно в горнодобывающих компаниях, судостроительных, для подачи сырья на склады или на производственные площадки нужны узкоколейные пути. Именно о таком подвиде и пойдет речь в текущем обзоре. Разберем рельс Р18 характеристики, плотность, размеры, ГОСТ, вес 1 метра и иные сопутствующие факторы, изучим параметры на схемах. И что важнее, точно определим предназначение и область эксплуатации такого полотна.
И если стандартное полотно по типу Р65 зачастую необходимо для обычной транспортировки грузов между муниципальными образованиями, то конкретно в горнодобывающих компаниях, судостроительных, для подачи сырья на склады или на производственные площадки нужны узкоколейные пути. Именно о таком подвиде и пойдет речь в текущем обзоре. Разберем рельс Р18 характеристики, плотность, размеры, ГОСТ, вес 1 метра и иные сопутствующие факторы, изучим параметры на схемах. И что важнее, точно определим предназначение и область эксплуатации такого полотна.
До изучения технических характеристик стоит пристально посмотреть на сферу эксплуатации этой продукции, чтобы понять, будет ли вариант эффективен конкретно в вашем случае. Железная дорога была изобретена уже очень давно. Но первые ее прототипы как раз и базировались на узкой колее. В те времена не было возможность развернуться производство полноценных путей на железобетонных шпалах, способных выдержать высокий трафик составов. Поэтому тяжелые рельсы, Р65, Р75 не были необходимыми. Интересно то, что с течением времени актуальность узкоколейных железнодорожных путей не снизилась. Напротив, если сложить все участки такого типа, их длину, то она не уступит широким путевым линиям в совокупности. Просто зачастую они скрыты от глаз.
Поэтому тяжелые рельсы, Р65, Р75 не были необходимыми. Интересно то, что с течением времени актуальность узкоколейных железнодорожных путей не снизилась. Напротив, если сложить все участки такого типа, их длину, то она не уступит широким путевым линиям в совокупности. Просто зачастую они скрыты от глаз.
Это линии сообщений, соединяющие между собой сырьевые базы, склады, площадки для производства, заводы. На основе такого вида строятся крановые мосты, опорные кран-балки, дороги прокладываются внутри шахт, рудников. Это часто вагонеточные пути, по которым также могут передвигаться специальный автоматизированный транспорт небольших габаритов. Но, помимо этого, это еще и подача сырья на различных типах конвейерного производства. Отсюда вытекает две основные группы узкоколейных путей.
https://promputsnab.ru/obshee-opisanie-rels.html
Учитывая, что рельс 18 размеры имеет скромные, проложенные участки дороги легко помещаются в производственных зданиях, так сказать, под крышей. Внутри ангаров, складов, цехов. Такие участки называются внутренними. К внешним же относятся все остальные начиная от распространенных вагонеточных путей, до кран-балок.
Внутри ангаров, складов, цехов. Такие участки называются внутренними. К внешним же относятся все остальные начиная от распространенных вагонеточных путей, до кран-балок.
Также стоит сразу уточнить, что Р18 – это только один из двух основных видов узкой колеи. Не меньшей актуальностью пользуется и Р24. Логично даже использовать оба варианта в условиях одного производства. Например, более широкие типы можно применять для внешнего сообщения, а узкие уже внутри самого производственного здания.
Также стоит понимать, что все типы с более крупным размерным рядом уже относятся к стандартному полотну, и не могут применяться в основе для тех же вагонеточных скреплений. Помните, что качество продукты опирается не только на размеры, но и на сталь, из которой производится продукция. Распространены три ключевых варианта, это мартеновская сталь, электросталь и конвертерная. Они обозначаются соответствующей маркировкой: М, Э, К.
Также продукция может иметь термическое упрочнение. Это, соответственно, способ создать более надежный товар.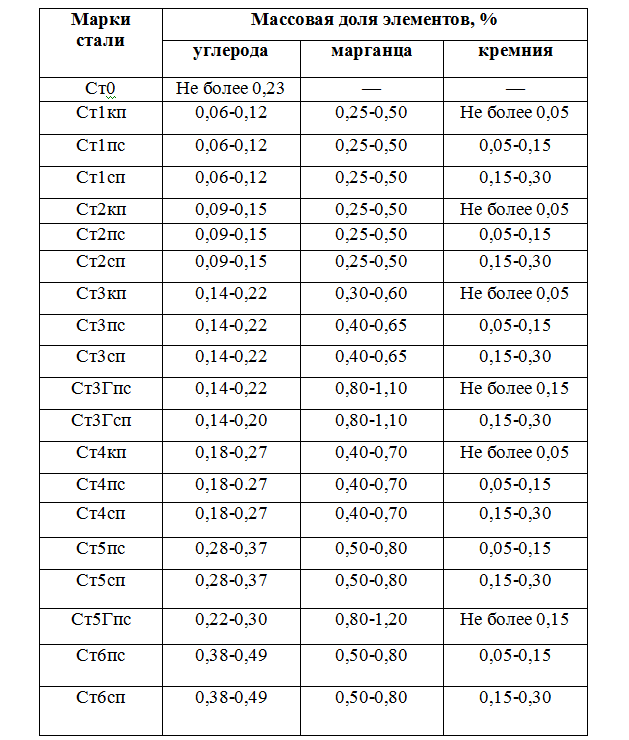 Но ценовая позиция тоже становится выше. В любом случае особенно если речь идет о крупной партии, важно подобрать надежного поставщика.
Но ценовая позиция тоже становится выше. В любом случае особенно если речь идет о крупной партии, важно подобрать надежного поставщика.
И отличным выбором станет «ПромПутьСнабжение». Очевидные плюсы сотрудничества в следующем:
Качественная продукция. Все товары проходят полноценные проверки на соответствие нормативным документам. Если говорить про рельс Р18 – ГОСТ 6368 82. Именно этот стандарт качества регламентирует все основные параметры эталонной продукции, выявляет нормы допустимых отклонений.
Огромный ассортимент. Логично предположить, что при укладке железнодорожного пути не получится обойтись только самим полотном. Понадобится масса метизов, предназначать которые нужно в качестве фиксирующих, предохраняющих, крепежных элементов. Это путевые болты, клеммы, гайки, прижимы, шайбы и требуемый для монтажа инструментарий. В ПромПутьСнабжение вы можете сразу приобрести все необходимое для выполнения работ по укладке. Не придется искать никакие дополнительные магазины, поставщиков.
 Один заказ – это товары для работы под ключ.
Один заказ – это товары для работы под ключ. Удобная ценовая политика. И конкретно сейчас стоит отметить, что как раз на узкоколейные рельсы действует специальная акция. Массовая распродажа со склада, которая позволит вам приобрести продукцию с впечатляющей скидкой. И это при том, что даже стандартный прейскурант в принципе весьма лоялен по сравнению со среднерыночной ценой. А так он становится еще более приветливым.
Доставка любыми удобными способами. Компания может взять все вопросы, связанные с транспортировкой на себя. Возможны различные варианты по желанию заказчику. Также исходя из объема поставок выбирается водное сообщение, железнодорожные пути или обычный автомобильный транспорт. И что важнее всего, главный критерий выбора – оперативность исполнения.
Теперь же перейдем к сугубо техническим параметрам, которые помогут точно разобраться в аспектах монтажа, транспортировки. Ведь без знания основных характеристик невозможно заблаговременно рассчитать корректный план-проект.
Рельсы R18, как и остальные их собратья, состоят из нескольких частей. И нижняя из них – это основание. Его еще принято называть «подошвой». Если разбирать вопрос на схеме, то эта плоскость обозначается буквой B. Именно к основанию подходят практически все крепежи. Поэтому зачастую это часть наиболее широкая из всех, ведь нагрузка будет приходиться постоянно.
В нашем случае «подошва» имеет ширину в 80 мм. Это, разумеется, сравнительно немного. Но для узкой колеи – это оптимальный выбор. Такая ширина позволит обеспечить надежную фиксацию полотна. И при этом не увеличить чрезмерно вес и ценовой параметр продукта.
Ширина верхней частиА это как раз те самые полозья, по которым и будут двигаться вагонетки или иные приспособления, доставляющие продукция, конвейерные механизмы и схожее. На схемах этот элемент можно найти под маркером b1. Ширина всего 40 мм, это ровно вдвое меньше, чем основание.
У этого элемента тоже есть свое обиходное название, «проезжая часть».
Показатель тоже является исключительно важным для расчета весовых параметров, транспортировки, хранения и, естественно, монтажа. Именно высота — наибольшее значение во всей конструкции (за исключением длины, которая может быть определена произвольно). На схеме обозначается, как H. И равняется по государственному стандарту качеству 90 мм.
Рельс 18: вес 1 метраНа этот вопрос нельзя ответить с точностью. Дело в том, что для расчета необходимо знать плотность стали, из которой произведен товар. Разумеется, существует всего три варианта, о которых мы уже сказали выше. При этом еще и актуальностью пользуется только один из них. Фактически в 99% подразумевается 7830 кг/м3. И исходя из этого показателя и необходимо проводить все расчетные операции.
Выявить этот параметр для целой линии проблематично, неясно какой длины продукция приобретена. Стандарт обычно – 8 м. Но при этом многие производители могут реализовывать и укороченные типы, а также и удлиненные. «ПромПутьСнабжение» сможет подобрать варианты длиной всего от 3 до 12 м. Поэтому если мы обсуждаем рельс Р 18 вес выявляется именно в погонном метре. И это ровно 17.96 кг.
«ПромПутьСнабжение» сможет подобрать варианты длиной всего от 3 до 12 м. Поэтому если мы обсуждаем рельс Р 18 вес выявляется именно в погонном метре. И это ровно 17.96 кг.
https://promputsnab.ru/relsy-p18-p24.html
Особенности изготовленияКак мы уже отмечали, хороший товар возможен только при качественном производстве. А значит, производственный аспект выходит чуть ли не на лидерскую позицию по важности. В государственном стандарте указаны рекомендации, но методики совершенствуются постоянно. И нормативная документация не может адаптироваться под каждую новинку технического прогресса, которая внедряется на заводах. Поэтому сам процесс производства с нюансами зачастую – это выбор производителя.
Разумеется, есть некоторые базовые аспекты, которые всегда имеют место, как:
Обязательная термическая обработка для уплотнения.
Строго перпендикулярные срезы на торцах. Логично, если будет отклонение по оси хоть на десятую долю градуса, то велик шанс, что при стыковке двух рельсов между собой возникнут серьезные сложности.
 В стандарте устанавливается допуск в 1 миллиметр, но не рекомендуется близко к нему подбираться.
В стандарте устанавливается допуск в 1 миллиметр, но не рекомендуется близко к нему подбираться. Отверстия для болтов должны размещаться равномерно, чтобы распределить нагрузку. При этом самый первый болт нельзя крепить ближе, чем в 46 миллиметрах от стыка. Иначе при интенсивной эксплуатации может возникнуть деформация стали.
Чтобы получить действительно качественную продукцию, необходимо грамотно выбрать поставщика. Компания «ПромПутьСнабжение» работает только по проверенным технологиям со строгим контролем производственного процесса. Поэтому любые отклонения вне границ допусков в товарах невозможны по определению. Все рельсы Р 18 ГОСТ 6368 82 вышедшие с производственных площадок – соответствуют не только указанному стандарту, но также и многочисленным международным нормативам.
МаркировкаС помощью тщательного изучения схемы, также самого изделия можно легко понять все его основные характеристики. Так внешний визуальный осмотр поможет определить каленую продукцию. Для этого достаточно посмотреть на рельсовую шейку. Там должна располагаться специальная поперечная полоса, которая явно указывает на то, что изделие было прокалено. Также на метизе стоит поискать специальное кольцо, вдавленное на небольшом углублении.
Для этого достаточно посмотреть на рельсовую шейку. Там должна располагаться специальная поперечная полоса, которая явно указывает на то, что изделие было прокалено. Также на метизе стоит поискать специальное кольцо, вдавленное на небольшом углублении.
На схеме же маркировка следующая:
B. Как мы уже отмечали выше, это ширина нижней части метиза. То есть той, что называется подошвой.
b1. Это уже размер верхней поверхности, по которой движется вагонетка или автоматизированный транспорт.
H. Общая высота всего изделия вместе с подошвой, поперечной и верхней частью.
S. Величина нейтральной оси.
M. Масса, как мы помним, она обозначается в погонных метрах. На некоторых чертежах размещается этот показатель общей линии в совокупности, но конкретно при разборе рельсы, обычно указывается только номинальные значения.
Даже самое точное описание металлического изделия не позволит со 100% точностью уяснить все необходимые аспекты. Для этих целей используются схемы, где наглядно изображаются все размерные ряды участков, изгибы, углы отклонений и иные элементы. Подобного рода изображения входят в общий пакет документации, которая будет нужной для укладки железнодорожных путей. Часть передает сам производитель в качестве сопутствующих документов, а другую — на месте формирует инженерная группа.
Для этих целей используются схемы, где наглядно изображаются все размерные ряды участков, изгибы, углы отклонений и иные элементы. Подобного рода изображения входят в общий пакет документации, которая будет нужной для укладки железнодорожных путей. Часть передает сам производитель в качестве сопутствующих документов, а другую — на месте формирует инженерная группа.
Теперь давайте более детально затронем области, в которых подразумевается эксплуатация Р18 ГОСТ 6368 82. А это, как мы уже поняли, узкоколейные железнодорожные пути.
Горнодобывающая промышленность. Соответственно, это рудники, шахты, подземные пути сообщений. В условиях ограниченного пространства нет никакой возможности развернуть полноценное строительство. Тяжелая техника просто не сможет проехать в шахту. Укладывать крупную железную дорогу, асфальтированную площадку невозможно. Также рельсовая решетка не может быть уложена в таких условиях, ведь ни кран, ни трактор просто не пройдет.
 Соответственно, единственным выходом становится ручной монтаж узкой колеи.
Соответственно, единственным выходом становится ручной монтаж узкой колеи. Размещение кран-балок и крановых мостов.
Поставка товаров, сырья, материалов от складов, сырьевых баз на место производства в условиях их расположения в относительной близости. В этом случае на первую позицию выходит уже экономия. Да, между точками можно развернуть практически любой вид сообщения. Но выгоднее становится узкая колея.
Транспортировка продукции при освоении целинных земель.
Военно-полевые дороги, которые распространены при формировании укреплений, фортов, новых гарнизонов.
Внутренние пути на предприятиях, складах, ангарах, сырьевых базах, цеха и заводах. Не секрет, что некоторые из обозначенных строений имеют очень крупный масштаб. И для быстрой передачи нескольких сотен товар сырья от одной точки к другой самым логичным выходом будет использование транспорта по типу вагонеток.
Развлекательная сфера. Все подвиды, так называемых «американских горок» также строятся на этом принципе.
 Как и колеи, предназначение для детских аттракционов, мини поездов.
Как и колеи, предназначение для детских аттракционов, мини поездов.
При этом фактически ни одно предприятие не имеет строгих ограничений на использование такого варианта. Перевозка сырья всегда может стать, как говорится, «на рельсы». Единственный разумный довод «против», если объем производства, а с ним и транспортировки сырьевых основ – ничтожно мал. Тогда укладка железной дороги не будет рентабельной. Также аргументов может стать временный характер производства. Ведь демонтировать конструкцию после исполнения задачи – это затратное мероприятие, которое не факт, что себя окупит.
Расчетные значения размеровРассмотрим все основные параметры в виде наглядной таблицы.
Тип | S поперечного сечения в квадратных сантиметрах | Дистанция от головки до центра | Сопротивление по верху | По низу | По грани | m |
Р 18 | 22. | 4.6 | 50.8 | 55.3 | 10.1 | 17.9 |
 8
8Изделия из стали нельзя назвать прихотливыми. Но они также при длительном хранении с неправильными условиями могут значительно снизить свой предельный эксплуатационный срок. Основным аспектом, который пагубно влияет на сталь, является климатические условия. Зачастую временно рельсы располагают на открытом воздухе. И даже если предварительно накрыть их тентом, скопления влаги все же может появиться. Чтобы предотвратить этот фактор, необходимо всегда размещать площадку хранения выше по уровню, относительно близлежащего рельефа местности.
ТранспортировкаВ этом моменте важно обратить свое внимание на два ключевых фактора. Первый заключается в значительном весе товара, если речь идет о крупной поставке. Нужно сразу рассчитать тот факт, что не каждый транспорт с этим справится. И второй момент – длина. Если это более 8 метров, да еще и несколько десятков или сотен тонн одновременно, то автомобильный транспорт сразу исключается. Логично будет использовать доставку железной дорогой или судном.
Если это более 8 метров, да еще и несколько десятков или сотен тонн одновременно, то автомобильный транспорт сразу исключается. Логично будет использовать доставку железной дорогой или судном.
Мы досконально разобрали рельсы Р 18. И теперь темных пятен не осталось. Остается лишь сверить конкретно ваши требования на производстве с условиями, при которых узкая колея становится актуальной. И если они соответствуют, то перейти к выбору поставщика.
Марка стали Р18
- Главная
- Информация
- Справочник марок стали
- Сталь инструментальная быстрорежущая
 org/ListItem”>Р18
org/ListItem”>Р18
Другие марки сталиУ8А4Х5МФСР6М5У10АР9К5Р6М5К5Ст40Х12МФСт456ХВ2С65ГСт505ХНМ60С2А40х134Х4ВМФС40х4Х5В2ФСCталь 30ХГСА9ХСШХ15ХВГСт20Ст35
| Марка: | Р18 |
| Класс: | Сталь инструментальная быстрорежущая |
| Используется для проката: |
Сортовой и фасонный прокат: ГОСТ 19265-73 , ГОСТ 2590-2006, ГОСТ 2591-2006 Калиброванный пруток: ГОСТ 19265-73 , ГОСТ 7417-75 Шлифованный пруток и серебрянка: ГОСТ 19265-73 , ГОСТ 14955-77 Лист толстый: ТУ 14-1-1408-75 Лист тонкий: ТУ 14-1-1706-76, ТУ 14-1-1408-75 Полоса: ГОСТ 19265-73 , ГОСТ 4405-75 Проволока: ТУ 14-1-1096-74 Поковки и кованые заготовка: ГОСТ 19265-73, ГОСТ 1133-71.  |
| Использование в промышленности: | Из стали Р18 изготовляют резцы, фрезы, сверла, долбяки, развертки, зенкеры, метчики, протяжки и другие режущие инструменты. |
| Твердость материала: | HB 10-1 = 255 МПа |
| Температура критических точек: | Ac1 = 820 , Ac3(Acm) = 860 , Ar3(Arcm) = 770 , Ar1 = 725 |
| Температура ковки, °С: | Начала 1200, конца 900. Охлаждение в колодцах при 750-800 °С |
| Обрабатываемость резанием: | При HB 212-228 Кυ тв. спл. = 0,6 и Кυ б.ст. = 0,3 |
| Свариваемость материала: | Сварка производится без подогрева и без последующей термообработки (без ограничений). Хорошая свариваемость при стыковой электросварке со сталями 45 и 40Х |
| Флокеночувствительность: | Не чувствительна |
| Склонность к отпускной хрупкости: | Малосклонна |
| Аналоги: | Р9К5, Р6М5, Р6М5К5 |
Описание Р18
Быстрорежущая сталь Р18 применяется для изготовления всех видов режущего инструмента при обработке углеродистых легированных конструкционных сталей с прочностью до 1000 МПа, от которых требуется сохранение режущих свойств при нагревании во время работы до 600°С.
Расшифровка
- Буква Р — указывает, что сталь быстрорежущая;
- Цифра 18 — указывает на наличие Вольфрама (W) и его средний % в стали (в данной стали значение Вольфрама 17 – 18,5%).
Химический состав стали Р18
| Химический элемент | % |
|---|---|
| Углерод (C) | 0,73 – 0,83 |
| Кремний (Si) | 0,2 – 0,5 |
| Марганец (Mn) | 0,2 – 0,5 |
| Никель (Ni) | до 0,6 |
| Фосфор (P) | до 0,03 |
| Хром (Cr) | 3,8 – 4,4 |
| Молибден (Mo) | до 1 |
| Вольфрам (W) | 17 – 18,5 |
| Ванадий (V) | 1 – 1,4 |
| Кобальт (Co) | до 0,5 |
| Сера (S) | до 0,03 |
| Медь (Cu) | до 0,25 |
| Железо (Fe) | ~73 |
Температура критических точек, °С
| Ac1 | Ac3 | Ar1 | Ar3 |
|---|---|---|---|
| 820 | 860 | 725 | 770 |
Физические свойства Р18
| Температура испытания, °С | 20 | 100 | 200 | 300 | 400 | 500 | 600 | 700 | 800 | 900 |
|---|---|---|---|---|---|---|---|---|---|---|
| Модуль нормальной упругости E, ГПа | 228 | 223 | 219 | 210 | 201 | 192 | 181 | – | – | – |
| Плотность, pn, кг/см3 | 8800 | – | – | – | – | – | – | – | – | – |
| Коэффициент теплопроводности Вт/(м ·°С) | – | 26 | 27 | 28 | 29 | 28 | 27 | 27 | – | – |
| Удельное электросопротивление (p, НОм · м) | 419 | 472 | 544 | 627 | 718 | 815 | 922 | 1037 | 1152 | 1173 |
| Коэффициент линейного расширения (a, 10-6 1/°С) | – | – | – | – | – | – | – | – | – | – |
| Удельная теплоемкость (С, Дж/(кг · °С)) | – | – | – | – | – | – | – | – | – | – |
Механические свойства Р18 в зависимости от температуры отпуска
| Температура отпуска, °С | 400 | 500 | 550 | 600 |
|---|---|---|---|---|
Закалка 1280 °С, масло. Отпуск трехкратный по 1 ч Отпуск трехкратный по 1 ч | ||||
| σв (МПа) | 1370 | 1470 | 2350 | 2210 |
| KCU (Дж / см2) | 23 | 19 | 17 | – |
| HRC ∂ (HB) | 61 | 63 | 66 | 65 |
Механические свойства стали Р18 в термообработанном состоянии при повышенных температурах
| Температура испытания, °С | 200 | 400 | 500 | 550 | 600 | 650 |
|---|---|---|---|---|---|---|
| σизг. (МПа) | 3570 (180) | 3730 (180) | 3290 (160) | 3060 (150) | 2430 (120) | 2180 (110) |
| HV | 815 (10) | 755 (10) | 712 (10) | 661 (10) | 615 (10) | 504 (10) |
| HRC ∂ (HB) | 64 | 62 | 60 | 58 | 56 | 51 |
Механические свойства стали Р18 в состоянии поставки (после отжига) при повышенных температурах
| Температура испытания, °С | 200 | 400 | 600 | 800 | 1000 | 1100 | 1200 |
|---|---|---|---|---|---|---|---|
| σ0,2 (МПа) | 450 (50) | 420 (40) | 300 (40) | 110 (20) | 90 (20) | – | 30 (10) |
| σв (МПа) | 830 (80) | 700 (70) | 480 (50) | 200 (20) | 100 (20) | – | 30 (10) |
| δ5 (%) | 13 (2) | 15 (2) | 31 (3) | 60 (5) | 42 (4) | – | 12 (3) |
| ψ % | 22 (4) | 22 (4) | 55 (6) | 70 (6) | 55 (6) | – | 25 (5) |
σсж. (МПа) (МПа) |
1050 (50) | 850 (50) | 620 (40) | 100 (20) | 50 (10) | – | 40 (10) |
| Τк (МПа) | 520 (30) | 450 (30) | 300 (20) | 100 (20) | 50 (10) | – | 40 (10) |
| KCU (Дж / см2) | – | – | – | – | 100 (10) | 130 (15) | 45 (5) |
| HB | 227 (6) | 210 (6) | 140 (6) | 30 (4) | 24 (4) | – | 4 (1) |
Р18 :: Металлические материалы: классификация и свойства
Сталь Р18 ГОСТ 19265-73
Массовая доля элемента, % | |||||||||||||
углерода | марганца | кремния | хрома | вольфрама | ванадия | кобальта | молибдена | никеля | меди | серы | фосфора | азота | ниобия |
не более | |||||||||||||
0,73– 0,83 | 0,20 – 0,50 | 0,20 – 0,50 | 3,80 – 4,40 | 17,00 – 18,50 | 1,00 – 1,40 | Не более 0,50 | Не более 1,00 | 0,6 | 0,25 | 0,030 | 0,030 | – | – |
Ас1 | Ас3 (Асm) | Ar3(Асm) | Мн | |
Температура критических точек, °С | 820 | 860 | 770 | 725 |
Температура испытания °С | ||||||||||
20 | 100 | 200 | 300 | 400 | 500 | 600 | 700 | 800 | 900 | |
Модуль нормальной упругости, Е, ГПа | 228 | 223 | 219 | 210 | 201 | 192 | 181 | – | – | – |
Модуль упругости при сдвиге кручением G, ГПа | 83 | – | – | – | – | – | – | – | – | – |
Плотность ρn, г/см3 | 8800 | – | – | – | – | – | – | – | – | – |
Коэффициент теплопроводности λ, Вт/(м * °С) | – | 26 | 27 | 28 | 29 | 28 | 27 | 27 | – | – |
Удельное элекросопротивление (ρ, Ном*м) | 419 | 472 | 544 | 627 | 718 | 815 | 922 | 1037 | 1152 | 1173 |
Твердость | Температура 0С | ||
после отжига | После закалки с отпуском, HRCэ (HRC), не менее | закалки | отпуска |
HB, не более | |||
255 | 63 (62) | 1270 | 560 |
Температура нагрева для
закалки и отпуска образцов должна соответствовать значениям, указанным в
таблице.
Охлаждение образцов после закалки проводят в масле.
Отпуск образцов проводят двух-, трехкратный, с выдержкой по 1 ч. и охлаждением на воздухе.
Время выдержки при нагреве устанавливают согласно черт. 1.
Толщина (диаметр) образца, мм
1. – для прямоугольных образцов
2. – для круглых образцов
Черт. 1.
Кривая зависимости твердости от температуры отпуска
Механические свойства стали в состоянии поставки (после отжига) при 20 °С
σ0,05 | σ0,2 | σВ | δ5 | ψ | σСЖ0,2 | σСЖ | ε, % | τК, МПа | ν, % | KCU, Дж/см2 |
МПа | % | Мпа | ||||||||
210 (5) | 510 (20) | 840 (30) | 8 (1) | 10 (1) | 520 (13) | 2600 (80) | 50 (1,5) | 560 (17) | 50 (0,7) | 19 (1) |
Механические свойства стали в термообработанном состоянии
σ0,05 | σВ | σСЖ0,2 | σСЖ | τК, | σизг | KCU, Дж/см2 |
МПа | ||||||
2480 (70) | 2150 (110) | 3060 (90) | 3820 (120) | 1880 (100) | 3000 (200) | 30 (3) |
Механические свойства стали в состоянии поставки (после отжига) при повышенных температурах
Температура испытания, °С | σ0,2 | σВ | δ5 | ψ | σсж | τК, | KCU Дж/см2 | НВ |
МПа | % | МПа | ||||||
200 | 450 (50) | 830 (80) | 13 (2) | 22 (4) | 1050 (50) | 520 (30) | – | 227 (6) |
400 | 420 (40) | 700 (70) | 15 (2) | 22 (4) | 850 (50) | 450 (30) | – | 210 (6) |
600 | 300 (40) | 480 (50) | 31 (3) | 55 (6) | 620 (20) | 300 (20) | – | 140 (6) |
800 | 110 (20) | 200 (20) | 60 (5) | 70 (6) | 100 (20) | 100 (20) | – | 30 (4) |
1000 | 90 (20) | 100 (20) | 42 (4) | 55 (6) | 50 (10) | 50 (10) | 100 (10) | 24 (4) |
1100 | – | – | – | – | – | – | 130 (15) | – |
1200 | 30 (10) | 30 (10) | 12 (3) | 25 (5) | 40 (10) | 40 (10) | 45 (5) | 4 (1) |
Механические свойства стали в термообработанном состоянии при повышенных температурах
Температура испытания, °С | σизг, МПа | HV | HRC |
200 | 3570 (180) | 815 (10) | 64 |
400 | 3730 (180) | 755 (10) | 62 |
500 | 3290 (160) | 712 (10) | 60 |
550 | 3060 (150) | 661 (10) | 58 |
600 | 2430 (120) | 615 (10) | 56 |
650 | 2180 (110) | 504 (10) | 51 |
Механические свойства в зависимости от температуры отпуска
Температура отпуска, °С | σВ МПа | KCU Дж/см2 | НRCЭ | |
Закалка 1280 °С, масло. |
| |||
400 | 1370 | 23 | 61 |
|
500 | 1470 | 19 | 63 |
|
550 | 2350 | 17 | 66 |
|
600 | 2210 | – | 65 |
|
 Отпуск трехкратный по 1 ч
Отпуск трехкратный по 1 чВязкость | Сопротивление износу | Шлифуемость | Красностойкость 59HRCэ при отпуске в течении 4ч, 0С | Особые свойства |
Хорошая | Хорошее | Повышенная | 620 | Пониженная
склонность к нагреву при закалке. |

Заменитель – сталь Р12
Температура ковки, °С:
начала 1200,
конца 900.
Охлаждение в колодцах при 750 – 800 °С.
Свариваемость – хорошая при стыковой электросварке со сталями 45 и 40Х.
Обрабатываемость резанием – при HB 212 – 228, Кυ тв. спл=0,6; Кυ б. ст=0,3.
Применение: Резцы, сверла фрезы, резьбовые фрезы, долбяки, развертки, зенкеры, метчики, протяжки для обработки конструкционных сталей с прочностью до 1000 МПа, от которых требуется сохранение режущих свойств при нагревании во время работы до 600 °С.
Сортамент:
горячекатаная круглого и квадратного сечений – ГОСТ 2590-88 и ГОСТ 2591-88;
кованая – ГОСТ 1133-71;
полосовая – ГОСТ 4405-75;
калиброванная – ГОСТ 7417-75;
сталь со специальной отделкой поверхности – ГОСТ 14955-77 диаметром от 1 до 25 мм включительно.
Исследование стали Р18, отлитой центробежно-вакуумным способом Текст научной статьи по специальности «Технологии материалов»
ИЗВЕСТИЯ
ТОМСКОГО ОРДЕНА ТРУДОВОГО КРАСНОГО ЗНАМЕНИ ПОЛИТЕХНИЧЕСКОГО
ИНСТИТУТА имени С. М. КИРОВА
М. КИРОВА
1970
Том 173
ИССЛЕДОВАНИЕ СТАЛИ Р18, ОТЛИТОЙ ЦЕНТРОБЕЖНО-ВАКУУМНЫМ СПОСОБОМ
А. Н. ДОБРОВИДОВ, А. И. СЛОСМАН
(Представлена научным семинаром кафедр металловедения, технологии металлов
и сварочного производства)
Метод центробежной разливки в вакууме позволяет объединить достоинства вакуумной плавки (получение металла строго заданного состава, чистого по газам и неметаллическим включениям, с высокими механическими и физическими свойствами) с преимуществами центробежного способа разливки (получение плотной отливки с хорошей структурой и чистой и точной поверхностью) [1]. Поэтому этот метод дает возможность изготовления отливок более высокого, качества по сравнению с отливками, полученными другими способами. В частности, он может быть применен для существенного повышения качества литого инструмента.
Для выяснения целесообразности применения центробежной отливки в вакууме при изготовлении литого режущего инструмента было проведено исследование быстрорежущей стали типа Р18, выплавленной и разлитой в различных условиях.
Плавка производилась в индукционной вакуумной печи типа ИВ-52. Печь была приспособлена для центробежно-вакуумной разливки: был изготовлен индуктор, рассчитанный на плавку шихты весом 4—6 кг, и в печь был вмонтирован механизм для центробежной отливки в кокиль [2]. Схема реконструированной печи дана на рис. 1.
В качестве материала тигля применялись электроплавленный магнезит, хромомагнезит, окись алюминия, двуокись циркония. В случае применения магнезита, хромомагнезита и окиси алюминия тигель изготавливался путем спекания смеси соответствующего состава вокруг графитового шаблона, нагреваемого током высокой частоты до температуры выше 2000°; при этом происходило спекание тигля с оплавлением его внутренней поверхности. Цирконовые тигли, изготовленные на заводе высокоогнеупорных изделий, вставлялись непосредственно в индуктор. Тигли из магнезита и хромомагнезита имели меньшую стойкость, чем тигли из окиси алюминия и двуокиси циркония. Загрязнения выплавляемого металла материалами футеровки при наших условиях плавки не наблюдалось. 2 мм рт. ст. Натекание, определенное для холодной печи, составляло около 15 мк/мин.
2 мм рт. ст. Натекание, определенное для холодной печи, составляло около 15 мк/мин.
Источником тока служил преобразователь типа ПВ-50/2500. Установка работала на следующем режиме:
ток возбуждения генератора 6—7 а,
напряжение на индукторе ток индуктора COS ф
400—500 в, 90—120 а, 0,7—0,9.
Процесс нагрева и плавки шихты продолжался 10—15 мин. Затем жидкий металл выдерживался в вакууме 3—10 мин. Во время выдержки происходило выделение газов из металла, всплывание и разложение неметаллических включений.
Для того, чтобы предотвратить «кипение» жидкого металла при его охлаждении, давление в печи перед разливкой повышалось до 40—50 мм рт. ст. путем введения в нее аргона. После введения аргона выключалось питание индуктора, включался мотор центробежной машины и производилась разливка. До 300—400° отливка охлаждалась в вакууме.
На рис. 2 показан кокиль, примененный в работе. Материал кокиля— малоуглеродистая сталь. Плавки отливались в различных условиях (табл. 1). При разливке в открытой печи качество поверхности отливок было хуже: имели место плены, подкорковые пузыри.
1). При разливке в открытой печи качество поверхности отливок было хуже: имели место плены, подкорковые пузыри.
Шихтой для плавок служили куски стали Р18 из отходов инструментального производства. Химический анализ показал, что при вакуумном переплаве угара хрома, вольфрама, ванадия не происходит. Объясняется это тем, что при температурах и давлениях, которые имеют место в печи во время выдержки, раскислительная способность углерода превосходит раскислительную способность других легирующих элементов [3]. Поэтому, если в расплаве имеется достаточное количество углерода (в частности, при выплавке быстрорежущей стали), то окисления легирующих элементов не происходит. За счет испарения легирующих элементов химический состав стали тоже не может заметно измениться, так как упругость паров этих элементов в условиях выдержки
4
Рис. 1. Установка для плавки и центробежной разливки в вакууме: 1 — кожух, 2 — механизм вращения кокиля» 3 — кокиль, 4 — воронка, 5 — тигель, 6 —■ крышка кожуха% 7 — крышка горловины, 8 — меха-
низм поворота установки
близка (или ниже) упругости пара железа [4]. Уменьшения содержания металлических легирующих элементов за счет взаимодействия со стенками тигля не наблюдается. Углерод в применяемых условиях плавки практически тоже не взаимодействует с футеровкой. Для существенного развития реакции взаимодействия углерода, растворенного в жидком металле, с тиглем необходимы более низкие давления и более длительные выдержки [5]. Угар углерода зависит от времени выдержки жидкого металла в вакууме и идет, в основном, за счет реакции с растворенным в металле кислородом. Для компенсации уменьшения содержания углерода в результате угара в состав шихты вводилось определенное количество специально изогтовленного белого чугуна, полученного науглероживанием быстрорежущей стали. Содержание вольфрама и других легирующих элементов в этом чугуне было такое же, как и в стали Р18, а содержание углерода колебалось от 4- до 4,5%.
Уменьшения содержания металлических легирующих элементов за счет взаимодействия со стенками тигля не наблюдается. Углерод в применяемых условиях плавки практически тоже не взаимодействует с футеровкой. Для существенного развития реакции взаимодействия углерода, растворенного в жидком металле, с тиглем необходимы более низкие давления и более длительные выдержки [5]. Угар углерода зависит от времени выдержки жидкого металла в вакууме и идет, в основном, за счет реакции с растворенным в металле кислородом. Для компенсации уменьшения содержания углерода в результате угара в состав шихты вводилось определенное количество специально изогтовленного белого чугуна, полученного науглероживанием быстрорежущей стали. Содержание вольфрама и других легирующих элементов в этом чугуне было такое же, как и в стали Р18, а содержание углерода колебалось от 4- до 4,5%.
После охлаждения отливки подвергались трехкратному отпуску при температуре 570°. Затем из них изготовлялись образцы для исследования структуры, теплостойкости, неметаллических включений, плотности, износостойкости, механических и физических свойств.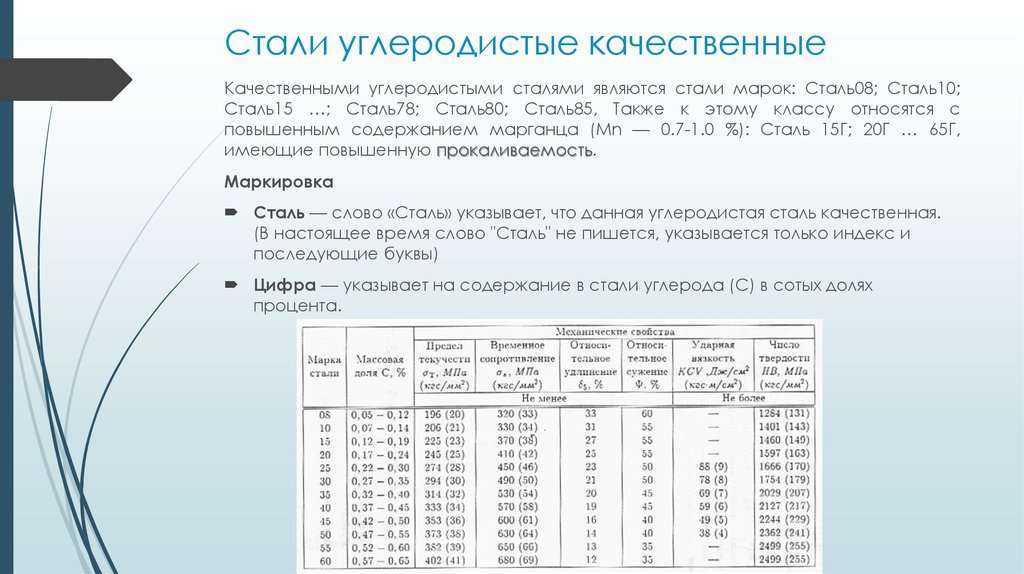
Для исследования макроструктуры образцы травили горячим 50%-ным раствором соляной кислоты в воде. Все отливки имели, в основном, плотную мелкозернистую структуру. В центральной части от-
Таблица 1
Условия выплавки и разливки исследованных плавок
№ плавок Давление во время плавки, мм рт. ст. Время выдержи, мин Давление во время разливки, мм рт. ст. Число оборотов кокиля, об ¡мин Температура разливки, °С
14 ю-2 5 50 300 1470—1500
15 5-КГ1 8 50 300 1540-1570
16 5-Ю”1 5 55 600 1470—1500
17 5- Ю-1 4 50 300 1470—1500
19 55 4 55 300 1470-1500
30 открытая печь 300 1470-1500
31 250 1540—1570
Примечания. 1. Давление в печи замерялось термопарным ва-кууметром ВТ-2. 2. Температура разливки определялась оптическим пирометром ОППИР-09.
,ливок наблюдались ‘незначительная пористость и несколько более грубое строение.
Микроструктура всех плавок типична для литой быстрорежущей стали. Она состоит из темной и светлой составляющих и эвтектики. Степень вакуума и число оборотов кокиля (в пределах от 200 до 800 об/мии) не оказали заметного влияния на размеры составляющих структуры. Очевидно, основными факторами, влияющими на формирование структуры, являются скорость охлаждения и химический состав отливок.
Степень вакуума и число оборотов кокиля (в пределах от 200 до 800 об/мии) не оказали заметного влияния на размеры составляющих структуры. Очевидно, основными факторами, влияющими на формирование структуры, являются скорость охлаждения и химический состав отливок.
В табл. 3 даны результаты измерения микротвердости отдельных составляющих плавок, сделанных в вакууме и в открытой печи.
При измерении твердости по Рок-веллу все отливки, независимо от условий выплавки и разливки, показали примерно одинаковую твердость. После литья они имели твердость 61—62 ЯС, а после трехкратного отпуска— 64—65 ЯС.
Теплостойкость определяли путем измерения твердости образцов в холодном состоянии. Образцы, подвергнутые после литья нормальному отпуску, нагревали до 600, 625 и 650° и выдерживали при этих температурах 4 часа. После охлаждения замеряли твердость. Результаты измерений показывают, что теплостойкость вакуумированной стали на 5—8 градусов выше, чем невакуумированной.
Таблица 3
Загрязненность неметаллическими включениями исследуемых плавок (по Герти)
№ плавок 14 15 16 17 19 30 31
Число пересечений на 1 см шлифа 13,9 14,7 17,1 14,4 19,2 42,0 35,5
Количественный металлографический анализ неметаллических включений, произведенный линейно-поперечным методом подсчета [6]л показал весьма значительное уменьшение загрязненности стали в результате вакуумного переплава. При этом методе исследования включений в окуляр вставляется стеклянная пластинка с пересекающимися под прямым углом линиями; нетравленый шлиф перемещается параллельно одной из них, и подсчитывается число включений, пересекаемых этой линией. Загрязненность выражается числом неметаллических включений, пересеченных линией при перемещении шлифа на 1 см. В работе ла каждом шлифе просматривалось по 10 см при увеличении 300, причем от каждой плавки бралось по 3 шлифа. Результаты анализа приведены в табл. 4.
При этом методе исследования включений в окуляр вставляется стеклянная пластинка с пересекающимися под прямым углом линиями; нетравленый шлиф перемещается параллельно одной из них, и подсчитывается число включений, пересекаемых этой линией. Загрязненность выражается числом неметаллических включений, пересеченных линией при перемещении шлифа на 1 см. В работе ла каждом шлифе просматривалось по 10 см при увеличении 300, причем от каждой плавки бралось по 3 шлифа. Результаты анализа приведены в табл. 4.
Кроме этого, для плавок 14 и 31 было определено содержание неметаллических включений в весовых процентах по методу, предложенному М. С. Ароновичем и И. М. Любарским [7]. Подсчет производился при увеличении 300. От каждой плавки брали по 3 шлифа и рассматривали по 60 полей на каждом из них. Получены следующие результаты: плавка 14 ( 10 мм рт. ст.) —0,0108% включений, плавка 31 (открытая печь)—0,0660% включений.
Таблица 2
Химический состав исследованных плавок
№ плавок Содержание элементов, %
с ы сг V
14 0,71 18,8 3,95 1,25
15 0,70 18,0 3,90 1,18
16 0,73 17,8 4,10 1,12
17 0,75 19,0 4,05 1,22
19 0,68 18,2 4,15 1,04
30 0,78 17,6 4,05 1,18
31 0,72 19,1 3,90 0,96
Качественный анализ неметаллических включений производился путем их рассматривания в обыкновенном свете в светлом и темном полях и в поляризованном свете при различных увеличениях. Анализ
Анализ
показал, что в вакууми-рованной стали неметаллические включения распределены более равномерно. Основную массу включений составляют алюмосиликатные стекла и окись алюминия. В стали, выплавленной в открытой печи, кроме того, обнаружены окислы других металлов (скорее всего железа, марганца, хрома, ванадия), а также сульфидов (очевидно сложных сульфидов FeS— —MnS).
Изучение влияния условий выплавки и разливки на плотность стали показало, что плотность повышается с увеличением степени вакуума при выплавке. Плотность определялась методом гидростатического взвешивания. Образцы взвешивались на воздухе и в дистиллированной воде. Увеличение плотности, наблюдаемое в результате вакуумного переплава, очевидно, объясняется уменьшением пористости и неметаллических включений.
Исследовалось также влияние степени вакуума в печи при выплавке на удельное электрическое сопротивление стали. Измерение проводилось на литых отпущенных и отожженных образцах. Данные об электросопротивлении образцов в литом отпущенном состоянии приведены на рис. 3. Очевидно, повышение электропроводности стали с повышением степени вакуума при ее выплавке связано с уменьшением в стали пор, неметаллических включений, газов, вредных примесей.
3. Очевидно, повышение электропроводности стали с повышением степени вакуума при ее выплавке связано с уменьшением в стали пор, неметаллических включений, газов, вредных примесей.
Выводы
1. При литье центробежно-вакуумным способом повышается качество поверхности отливок.
2. При плавке в вакууме быстрорежущей стали не происходит угара легирующих элементов. Это позволяет уменьшить пределы колебаний химического состава выплавляемой стали, а также дает экономию легирующих элементов.
3. Твердость быстрорежущей стали после переплава в вакууме не меняется; теплостойкость стали повышается на 5—8 градусов.
4. В результате вакуумного переплава в несколько раз уменьшается количество неметаллических включений, а оставшиеся включения распределяются более равномерно.
5. Центробежно-вакуумная разливка улучшает физические свойства стали (плотность, электропроводность).
6. Технических затруднений отливка в вакууме не представляет.
ЛИТЕРАТУРА
1. Д. Г. Люб едкий. Производство стали и отливок в вакууме. Машгиз. 1959.
Люб едкий. Производство стали и отливок в вакууме. Машгиз. 1959.
2. А. И. Слосман. Изв. ТПИ, т. 139. 1965.
3. F H and ers, H. Knüppel. К. Brotzmann Stahl und Eisen, 76, N 26, 1956.
4. E. С. Калинников. Вакуум в производстве стали. Металлургиздат. 1962.
5. H. A. Wrieght, J. С h i р m а п. Metallurgia, IX, 1195, 1956.
6. G. V. Hardy and R. T. Al Is о p. Journal of the Iron and Steel Inst, v. 195, part 3, 1960.
7. M. С. Дроков и ч, И. M. Л ю б a p с к и й. Металлург, № 9. 1936.
Ьаб/tefJue при плавке, мм рт с/п
Рис. 3
Ингибирование антисмысловой РНК экспрессии фосфопротеина p18 устраняет трансформированный фенотип лейкемических клеток
. 1996 15 марта; 56 (6): 1445-50.
С Джеха 1 , X N Luo, M Beran, H Kantarjian, GF Atweh
принадлежность
- 1 Отделение педиатрии, Онкологический центр доктора медицины Андерсона, Хьюстон, Техас 77030, США.
- PMID: 8640838
S Джеха и др. Рак Рез. .
. 1996 15 марта; 56 (6): 1445-50.
Авторы
С Джеха 1 , X N Luo, M Beran, H Kantarjian, GF Atweh
принадлежность
- 1 Отделение педиатрии, Онкологический центр доктора медицины Андерсона, Хьюстон, Техас 77030, США.
- PMID: 8640838
Абстрактный
Фосфопротеин р18 первоначально был идентифицирован на основании очень высокого уровня его экспрессии в лейкемических клетках разных линий. Изменения уровня накопления и фосфорилирования p18, связанные с индукцией дифференцировки лейкемических клеток, предполагают потенциальную роль этого фосфопротеина в клеточной пролиферации и дифференцировке и, возможно, в злокачественной трансформации. Недавние исследования показали, что p18 играет важную роль в развитии клеточного цикла, выступая в качестве субстрата для киназы p34(cdc2). Эти исследования показали, что ингибирование экспрессии p18 в лейкемических клетках приводит к замедлению роста и накоплению клеток в G(2)-M. В этом исследовании мы изучаем потенциальную роль p18 в клеточной трансформации, исследуя влияние ингибирования экспрессии p18 на злокачественный фенотип клеток эритролейкемии K562. Эти исследования показывают, что антисмысловое ингибирование экспрессии p18 в лейкемических клетках приводит к остановке роста при более низкой плотности насыщения, потере независимости от сыворотки и потере независимого от якоря роста in vitro. Кроме того, ингибирование экспрессии p18 приводит к заметному ингибированию онкогенности лейкозных клеток in vivo в мышиной модели с тяжелым комбинированным иммунодефицитом.
Изменения уровня накопления и фосфорилирования p18, связанные с индукцией дифференцировки лейкемических клеток, предполагают потенциальную роль этого фосфопротеина в клеточной пролиферации и дифференцировке и, возможно, в злокачественной трансформации. Недавние исследования показали, что p18 играет важную роль в развитии клеточного цикла, выступая в качестве субстрата для киназы p34(cdc2). Эти исследования показали, что ингибирование экспрессии p18 в лейкемических клетках приводит к замедлению роста и накоплению клеток в G(2)-M. В этом исследовании мы изучаем потенциальную роль p18 в клеточной трансформации, исследуя влияние ингибирования экспрессии p18 на злокачественный фенотип клеток эритролейкемии K562. Эти исследования показывают, что антисмысловое ингибирование экспрессии p18 в лейкемических клетках приводит к остановке роста при более низкой плотности насыщения, потере независимости от сыворотки и потере независимого от якоря роста in vitro. Кроме того, ингибирование экспрессии p18 приводит к заметному ингибированию онкогенности лейкозных клеток in vivo в мышиной модели с тяжелым комбинированным иммунодефицитом. Эти исследования демонстрируют, что высокий уровень экспрессии р18 в лейкемических клетках необходим для поддержания трансформированного фенотипа, и позволяют предположить, что р18 является потенциальной мишенью для противолейкозных вмешательств.
Эти исследования демонстрируют, что высокий уровень экспрессии р18 в лейкемических клетках необходим для поддержания трансформированного фенотипа, и позволяют предположить, что р18 является потенциальной мишенью для противолейкозных вмешательств.
Похожие статьи
Регуляция фосфопротеина р18 в лейкемических клетках. Фосфорилирование клеточного цикла регулируется киназой p34cdc2.
Луо XN, Мукерджи Б., Феррари А., Мистри С., Атвех Г.Ф. Луо XN и др. Дж. Биол. Хим. 1994 г., 8 апреля; 269(14):10312-8. Дж. Биол. Хим. 1994. PMID: 8144611
Транскрипционная регуляция фосфопротеина p18 во время моноцитарной дифференцировки U937 лейкозных клеток.
Мистри С., Луо XN, Атвех Г.Ф. Мистри С. и др. Cell Мол Биол Res.
 1995;41(2):103-10.
Cell Мол Биол Res. 1995.
PMID: 8581061
1995;41(2):103-10.
Cell Мол Биол Res. 1995.
PMID: 8581061Влияние ингибирования статмина на митотическое веретено.
Янку С., Мистри С.Дж., Аркин С., Валленштейн С., Атвех Г.Ф. Янку С. и др. Дж. Клеточные науки. 2001 март; 114 (часть 5): 909-16. doi: 10.1242/jcs.114.5.909. Дж. Клеточные науки. 2001. PMID: 11181174
Экспрессия статмина в иммортализованных и трансформированных онкогеном клетках.
Мистри С.Дж., Атве Г.Ф. Мистри С.Дж. и соавт. Противораковый Рез. 1999 г., январь-февраль; 19(1A):573-7. Противораковый Рез. 1999. PMID: 10226601
Роль статмина в регуляции митотического веретена: потенциальное применение в терапии рака.

Мистри С.Дж., Атве Г.Ф. Мистри С.Дж. и соавт. Гора Синай J Med. 2002 г., октябрь; 69 (5): 299–304. Гора Синай J Med. 2002. PMID: 12415323 Обзор.
Посмотреть все похожие статьи
Цитируется
AD80, мультикиназный ингибитор, проявляет противоопухолевое действие в клеточных моделях острого лейкоза, нацеленное на ось PI3K/STMN1.
Карлос ДЖАЭГ, Лима К., Коста-Лотуфо Л.В., Лейтао А., Мачадо-Нето Д.А. Карлос ЯЭГ и др. Инвестируйте в новые лекарства. 2021 авг; 39 (4): 1139-1149. doi: 10.1007/s10637-021-01066-w. Epub 2021 21 января. Инвестируйте в новые лекарства. 2021. PMID: 33475938
Ориентация на митотические пути для лечения рака, связанного с эндокринной системой.

Агарвал С., Варма Д. Агарвал С. и др. Endocr Relat Рак. 2017 сен;24(9)):T65-T82. DOI: 10.1530/ERC-17-0080. Epub 2017 14 июня. Endocr Relat Рак. 2017. PMID: 28615236 Бесплатная статья ЧВК. Обзор.
Экспрессия статмина 1 в неоплазмах плазматических клеток.
Мачадо-Нето Х.А., Периколь Ф.В., Коста Ф.Ф., Трайна Ф., Олалла Саад СТ. Мачадо-Нето Дж. А. и соавт. Rev Bras Hematol Hemoter. 2017 г., апрель-июнь; 39(2):183-185. doi: 10.1016/j.bjhh.2017.02.005. Epub 2017 11 марта. Rev Bras Hematol Hemoter. 2017. PMID: 28577660 Бесплатная статья ЧВК. Аннотация недоступна.
Ингибирование статмина-1 усиливает индуцированный руксолитинибом апоптоз в клетках JAK2V617F.
Мачадо-Нето Х.
 А., де Мело Кампос П., Фаваро П., Лазарини М., да Силва Сантос Дуарте А., Лоранд-Метце И., Коста Ф.Ф., Саад С.Т., Трайна Ф.
Мачадо-Нето Дж. А. и соавт.
Онкотаргет. 2015 6 октября; 6 (30): 29573-84. doi: 10.18632/oncotarget.4998.
Онкотаргет. 2015.
PMID: 26356819
Бесплатная статья ЧВК.
А., де Мело Кампос П., Фаваро П., Лазарини М., да Силва Сантос Дуарте А., Лоранд-Метце И., Коста Ф.Ф., Саад С.Т., Трайна Ф.
Мачадо-Нето Дж. А. и соавт.
Онкотаргет. 2015 6 октября; 6 (30): 29573-84. doi: 10.18632/oncotarget.4998.
Онкотаргет. 2015.
PMID: 26356819
Бесплатная статья ЧВК.SEPTIN2 и STATHMIN регулируют CD99-опосредованную клеточную дифференцировку при лимфоме Ходжкина.
Цзянь В., Чжун Л., Вэнь Дж., Тан Ю., Цю Б., Ву З., Ян Дж., Чжоу С., Чжао Т. Цзянь В. и др. ПЛОС Один. 22 мая 2015 г .; 10 (5): e0127568. doi: 10.1371/journal.pone.0127568. Электронная коллекция 2015. ПЛОС Один. 2015. PMID: 26000982 Бесплатная статья ЧВК.
Просмотреть все статьи “Цитируется по”
Типы публикаций
термины MeSH
- HL-42919/HL/NHLBI NIH HHS/США
- M01-RR00071/RR/NCRR NIH HHS/США
- Разделенный вид
- Содержание статьи
- Рисунки и таблицы
- видео
- Аудио
- Дополнительные данные
Цитировать
Cite
Shue Huang, Zhe Huang, Gregory Shearer, Shanshan Li, Shuohua Chen, Shouling Wu, Xiang Gao, Употребление алкоголя и возникновение инфаркта миокарда: роль холестерина липопротеинов высокой плотности (P18-020-19), Current Developments in Nutrition , Volume 3, Issue Supplement_1, June 2019, nzz039.
 P18–020–19, https://doi.org/10.1093/cdn/nzz039.P18-020-19
P18–020–19, https://doi.org/10.1093/cdn/nzz039.P18-020-19Выберите формат Выберите format.ris (Mendeley, Papers, Zotero).enw (EndNote).bibtex (BibTex).txt (Medlars, RefWorks)
Закрыть
Разрешения
- Электронная почта
- Твиттер
- Фейсбук
- Подробнее
Последний
Самые читаемые
Самые цитируемые
- Разделенный экран
- Просмотры
- Содержание артикула
- Рисунки и таблицы
- Видео
- Аудио
- Дополнительные данные
- Экспертная оценка
- Делиться
- Твиттер
- Инструменты
Получить разрешения
Иконка Цитировать Цитировать
- Поиск по сайту
- Значок версии статьи Версии
- Версия записи 1 19 марта96
- Рис (Зотеро)
- Менеджер ссылок
- EasyBib
- Подставки для книг
- Менделей
- Бумаги
- КонецПримечание
- РефВоркс
- Бибтекс
Zhang H, Hu X, Tse J, Tilahun F, Qiu M, Chen L . Спонтанное образование и регрессия лимфатических сосудов в роговице мыши. Invest Ophthalmol Vis Sci 2011; 52 :334–8.
Артикул Google ученый
Дэвидсон А.Дж. Развитие почек мыши. 2008. (http://www.ncbi.nlm.nih.gov/pubmed/20614633.)
Saint-Geniez M, D’Amore PA . Развитие и патология гиалоидных, хориоидальных и ретинальных сосудов. Int J Dev Biol 2004; 48 :1045–58.
Артикул Google ученый
Келлер Г., Циммер Г., Молл Г., Ритц Э., Аманн К. . Число нефронов у больных с первичной артериальной гипертензией. N Engl J Med 2003; 348 :101–8.
Артикул Google ученый
Занди-Нежад К.
 , Люкс В.А., Бреннер Б.М. Взрослая гипертензия и заболевание почек: роль фетального программирования. Гипертония 2006; 47 :502–8.
, Люкс В.А., Бреннер Б.М. Взрослая гипертензия и заболевание почек: роль фетального программирования. Гипертония 2006; 47 :502–8.КАС Статья Google ученый
Бертрам Дж.Ф., Дуглас-Дентон Р.Н., Диуф Б., Хьюсон М.Д., Хой В.Е. Число нефронов человека: последствия для здоровья и болезней. Педиатр Нефрол 2011; 26 :1529–33.
Артикул Google ученый
Ларссон Л., Аперия А., Уилтон П. . Влияние нормального развития на компенсаторный рост почек. Kidney Int 1980; 18 : 29–35.
КАС Статья Google ученый
Томат А.Л., Инсерра Ф., Вейрас Л. и др. Умеренное ограничение цинка во время внутриутробного и постнатального роста крыс: влияние на артериальное давление взрослых и почки. Am J Physiol Regul Integr Comp Physiol 2008; 295 :R543–9.
КАС Статья Google ученый
Влодек М.Е., Мибус А., Тан А., Зибель А.Л., Оуэнс Дж.А., Мориц К.М. Нормальная лактационная среда восстанавливает запас нефронов и предотвращает гипертензию после плацентарной рестрикции у крыс. J Am Soc Nephrol 2007; 18 :1688–96.
КАС Статья Google ученый
Шредер М.Ф., Ниенгард Дж.Р., Реммерс Ф., ван Вейк Дж.А., Делемарре-ван де Ваал Х.А. Постнатальное ограничение пищи у крыс как модель низкого запаса нефронов. Am J Physiol Renal Physiol 2006; 291 :F1104–7.
КАС Статья Google ученый
Ниенгаард младший . Стереологические методы и их применение в исследовании почек. J Am Soc Nephrol 1999; 10 :11:00–23.
КАС Google ученый
Каллен-МакИвен Л.
А., Армитаж Дж.А., Ниенгард Дж.Р., Мориц К.М., Бертрам Дж.Ф. Основанный на дизайне метод оценки количества клубочков в развивающейся почке. Am J Physiol Renal Physiol 2011; 300 :F1448–53.КАС Статья Google ученый
Вейбель Э.Р., Гомес Д.М. Принцип подсчета тканевых структур на случайных срезах. J Appl Physiol 1962; 17 :343–8.
КАС Статья Google ученый
Адамчак М., Гросс М.Л., Аманн К., Ритц Э. . Реверсия гломерулярных поражений включает скоординированную реструктуризацию гломерулярной микрососудистой сети. J Am Soc Нефрол 2004; 15 :3063–72.
Артикул Google ученый
Weibel ER . Численная плотность: форма и размер частиц. В: Weibel ER, изд. Стереологические методы Vol. 2: Теоретические основы.
Лондон: Academic Press, 1980: 140–74.Сэмюэл Т., Хой В.Е., Дуглас-Дентон Р., Хьюсон М.Д., Бертрам Дж.Ф. Применимость коэффициента распределения клубочков по размерам для оценки объема клубочков человека: пересмотр метода Вейбеля и Гомеса. Ж Анат 2007; 210 : 578–82.
Артикул Google ученый
Вульф КАК . Ангиопоэтины: факторы роста сосудов ищут роли в клубочках. Curr Opin Nephrol Hypertens 2010; 19 :20–5.
КАС Статья Google ученый
Огюстен Х.Г., Кох Г.Ю., Терстон Г., Алитало К. . Контроль морфогенеза сосудов и гомеостаза через систему ангиопоэтин-Tie. Nat Rev Mol Cell Biol 2009; 10 : 165–77.
КАС Статья Google ученый
Мирочник Ю., Квятек А.
, Вольперт О.В. Тромбоспондин и апоптоз: молекулярные механизмы и использование для разработки дополнительных методов лечения. Curr Drug Targets 2008; 9 :851–62.КАС Статья Google ученый
Кэмпбелл Н., Гринуэй Дж., Хенкин Дж., Петрик Дж. ABT-898 индуцирует регрессию опухоли и продлевает выживаемость в мышиной модели эпителиального рака яичников. Мол Рак Ther 2011; 10 : 1876–85.
КАС Статья Google ученый
Соломон С . Изменения числа нефронов, длины проксимальных канальцев и скорости клубочковой фильтрации поверхностных нефронов у крыс. J Physiol (Лондон) 1977; 272 : 573–89.
КАС Статья Google ученый
Хартман Х.А., Лай Х.Л., Паттерсон Л.Т. Прекращение почечного морфогенеза у мышей.
Dev Biol 2007; 310 : 379–87.КАС Статья Google ученый
Шевалье Р.Л., Торнхилл Б.А., Чанг А.И., Каша Ф., Лакей А. . Восстановление после устранения обструкции мочеточников у крыс: связь с нефрогенезом. Kidney Int 2002; 61 :2033–43.
Артикул Google ученый
Ниенгард младший . Количество и размеры гломерулярных капилляров крысы в норме и после нефрэктомии. Kidney Int 1993; 43 :1049–57.
КАС Статья Google ученый
Де Шпигелере В., Корнилли П., Симоенс П., Ван ден Брок В. . Иммуногистохимическое определение ангиопоэтинов при развитии метанефральной почки свиньи. Acta Histochem 2011; 113 :585–90.
КАС Статья Google ученый
Юань Х.
Т., Сури С., Лэндон Д.Н., Янкопулос Г.Д., Вульф А.С. Ангиопоэтин-2 является сайт-специфическим фактором дифференцировки почечной сосудистой сети мышей. J Am Soc Нефрол 2000; 11 :1055–66.КАС пабмед Google ученый
Юань Х.Т., Сури С., Янкопулос Г.Д., Вульф А.С. Экспрессия ангиопоэтина-1, ангиопоэтина-2 и тирозинкиназы рецептора Tie-2 во время созревания почек мыши. J Am Soc Nephrol 1999; 10 :1722–36.
КАС пабмед Google ученый
Дэвис Б., Деи Кас А., Лонг Д.А. и др. Специфическая для подоцитов экспрессия ангиопоэтина-2 вызывает протеинурию и апоптоз клубочкового эндотелия. J Am Soc Nephrol 2007; 18 :2320–9.
КАС Статья Google ученый
Ируэла-Ариспе М.Л., Лиска Д.Дж., Сейдж Э.
1
3131вещества
Грантовая поддержка
Потребление алкоголя и инфаркт миокарда: роль холестерина липопротеинов высокой плотности (P18-020-19) | Текущие разработки в области питания
Фильтр поиска панели навигации Текущие разработки в области питанияЭтот выпускЖурналы ASNДиетика и питаниеКнигиЖурналыOxford Academic Термин поиска мобильного микросайта
Закрыть
Фильтр поиска панели навигации Текущие разработки в области питанияЭтот выпускЖурналы ASNДиетика и питаниеКнигиЖурналыOxford Academic Термин поиска на микросайте
Расширенный поиск
Журнальная статья
Шу Хуанг,
Шу Хуанг
Ищите другие работы этого автора на:
Оксфордский академический
Google ученый
Чжэ Хуан,
Чжэ Хуан
Ищите другие работы этого автора на:
Оксфордский академический
Google ученый
Грегори Ширер,
Грегори Ширер
Ищите другие работы этого автора на:
Оксфордский академический
Google ученый
Шаньшань Ли,
Шаньшань Ли
Ищите другие работы этого автора на:
Оксфордский академический
Google ученый
Шуохуа Чен,
Шуохуа Чен
Ищите другие работы этого автора на:
Оксфордский академический
Google ученый
Шоулин Ву,
Шоулинг Ву
Ищите другие работы этого автора на:
Оксфордский академический
Google ученый
Сян Гао
Сян Гао
Ищите другие работы этого автора на:
Оксфордский академический
Google ученый
Current Developments in Nutrition , Volume 3, Issue Supplement_1, June 2019, nzz039. P18-020-19, https://doi.org/10.1093/cdn/nzz039.P18-020-19
P18-020-19, https://doi.org/10.1093/cdn/nzz039.P18-020-19
Опубликовано:
13 июня 2019 г.
Фильтр поиска панели навигации Текущие разработки в области питанияЭтот выпускЖурналы ASNДиетика и питаниеКнигиЖурналыOxford Academic Термин поиска мобильного микросайта
Закрыть
Фильтр поиска панели навигации Текущие разработки в области питанияЭтот выпускЖурналы ASNДиетика и питаниеКнигиЖурналыOxford Academic Термин поиска на микросайте
Расширенный поиск
Цели
Целью данного исследования является проверка гипотезы о том, что более низкий риск инфаркта миокарда (ИМ), связанный с употреблением алкоголя, обусловлен его влиянием на повышение уровня холестерина липопротеинов высокой плотности (ЛПВП).
Методы
В это исследование были включены 81 827 китайских мужчин и женщин (средний возраст: 51 ± 12 лет) из исследования Kailuan, у которых не было сердечно-сосудистых заболеваний в 2006 г. (исходно) и которые наблюдались до декабря 2016 г. Потребление алкоголя оценивали с помощью анкеты и измеряли концентрацию холестерина ЛПВП. Инцидент ИМ при последующем наблюдении был первым случаем ИМ, подтвержденным просмотром медицинской документации. Многопараметрическая регрессия Кокса использовалась для моделирования связи между привычным потреблением алкоголя и рисками ИМ с поправкой на потенциальные коварианты, включая возраст, пол, образование, ежемесячный доход, профессию, статус курения, физическую активность, индекс массы тела, окружность талии, гипертонию, диабет. и общий холестерин. Опосредованный эффект холестерина ЛПВП оценивали с использованием причинно-опосредованного анализа (макрос SAS).
Результаты
В среднем за 9,6 лет наблюдения мы задокументировали 1095 инцидентов. Скорректированное отношение рисков (ОР) для ИМ составило 0,64 (95% доверительный интервал (ДИ), 0,54–0,76) для лиц, употребляющих алкоголь в настоящее время, по сравнению с теми, кто никогда не употреблял алкоголь, или ранее. Соотношение изменилось очень незначительно, до 0,65 (95% ДИ, 0,55–0,76) после дальнейшей корректировки концентрации холестерина ЛПВП. Медиационный анализ показал, что концентрации холестерина ЛПВП опосредовали небольшую, незначительную долю (1,8%) влияния алкоголя на ИМ.
Скорректированное отношение рисков (ОР) для ИМ составило 0,64 (95% доверительный интервал (ДИ), 0,54–0,76) для лиц, употребляющих алкоголь в настоящее время, по сравнению с теми, кто никогда не употреблял алкоголь, или ранее. Соотношение изменилось очень незначительно, до 0,65 (95% ДИ, 0,55–0,76) после дальнейшей корректировки концентрации холестерина ЛПВП. Медиационный анализ показал, что концентрации холестерина ЛПВП опосредовали небольшую, незначительную долю (1,8%) влияния алкоголя на ИМ.
Выводы
Потребление алкоголя ассоциировалось с более низким риском заболеваемости ИМ в большой китайской когорте. Наши результаты показывают, что польза, связанная с умеренным потреблением алкоголя, не связана с влиянием алкоголя на холестерин ЛПВП.
Источники финансирования
Начальный грант Колледжа здравоохранения и человеческого развития штата Пенсильвания и Программа начальных грантов Penn State CyberScience.
Этот контент доступен только в формате PDF.
Авторское право © Американское общество питания, 2019 г.
Авторское право © Американское общество питания, 2019 г. Скачать все слайды
Реклама
Цитаты
Альтметрика
Дополнительная информация о метриках
Оповещения по электронной почте
Оповещение об активности статьи
Предварительные уведомления о статьях
Получайте эксклюзивные предложения и обновления от Oxford Academic
Ссылки на статьи по номеру
Местные продукты питания для решения проблемы недоедания: исследование диеты и статуса питания женщин в сообществе коренных народов племен мунда в Джаркханде, Индия
Влияние вмешательств в области питания на снижение анемии у женщин репродуктивного возраста в странах с низким и средним уровнем дохода – мета-обзор
Холин, докозагексаеновая кислота и диарейные заболевания, связанные с задержкой роста, в исследовании случай-контроль
Оценка разнообразных предпочтений и ожиданий фермеров в отношении адаптации мер по обеспечению продовольственной безопасности и питания на юго-востоке Мадагаскара
Окружность средней части верхней части руки новорожденного определяет детей с низким весом при рождении и уязвимых детей: вторичный анализ
Реклама
Предикторы низкой минеральной плотности костной ткани в когорте больных ирландским муковисцидозом (CF) | SFEBES2013 | Общество эндокринологии BES 2013
Доступные для поиска тезисы презентаций на ключевых конференциях по эндокринологии
ISSN 1470-3947 (печать) | ISSN 1479-6848 (онлайн)
Поиск Выпуски/Конференции Цитировать О Наши услуги Политики Контакт Отказ от ответственности
Endocrine Abstracts (2013) 31 P18
СФЭБЭС2013 Стендовые презентации Кость (34 тезисов)
Том МакЭнери
1, , Найджел Глинн 1, , Седрик Гунаратнам 2, , Ноэль МакЭлвани 2, , Диармуид Смит 1, & Клэр МакГенри 1,518 просмотров
Принадлежности автора Посмотреть электронную постер Скачать электронный постер
1 Отделение эндокринологии, больница Бомонт, Дублин, Ирландия; 2 Отделение респираторной медицины, больница Бомонт, Дублин, Ирландия; 3 Королевский колледж хирургов Ирландии, Дублин, Ирландия.
Увеличение ожидаемой продолжительности жизни у пациентов с муковисцидозом привело к возникновению новых проблем в их лечении. Остеопороз при МВ связан со значительной заболеваемостью и является критерием исключения для трансплантации легких. Последние данные свидетельствуют о связи между дисгликемией и низкой минеральной плотностью костей (МПКТ). Мы стремились определить предикторы потери костной массы в когорте пациентов с муковисцидозом, посещающих третичный специализированный центр, и, в частности, связана ли дисгликемия с низкой МПК.
Мы провели ретроспективный обзор пациентов, включенных в нашу базу данных по муковисцидозу в больницах. Записанные данные включали демографические и антропометрические характеристики пациентов, биохимию, МПК, измеренную с помощью двухэнергетической рентгеновской абсорбиметрии (DXA), функцию легких, назначенные лекарства и количество госпитализаций. Влияние характеристик пациентов на МПК анализировали с помощью критерия х 2 для дискретных переменных и критерия Стьюдента t для непрерывных переменных. Корреляция Спирмена между переменными пациента и 9Было подсчитано 0443 Z баллов.
Корреляция Спирмена между переменными пациента и 9Было подсчитано 0443 Z баллов.
Полные данные были доступны для 92 пациентов. Средний возраст составил 25±5 лет, ИМТ 20,9±4 кг/м 2 . Шестьдесят три пациента (68%) имели Z -балл ≤ -1, из которых 17 имели Z -балл ≤ -2,5. 55 имели нормальную толерантность к глюкозе, 12 имели нарушенную толерантность к глюкозе и 29 больных диабетом, связанным с муковисцидозом, с HbA1c 5,4±0,5, 5,9±0,4 и 7,7±1,8% и Z -баллами -1,4±1,2, -1,4±1,0 и -1,4±1,5 соответственно. Нижний Z -баллы были связаны с плохой функцией легких, низкой массой тела и более высокими показателями госпитализации и использования антибиотиков, но не с дисгликемией.
Несмотря на недавние данные, свидетельствующие о связи между дисгликемией и потерей костной массы при МВ, мы не обнаружили такой связи в нашей ирландской когорте. Общая тяжесть заболевания, а не дисгликемия, является лучшим предиктором потери костной массы при МВ.
DOI: 10.1530/endoabs.31.P18
Том 31Пред. Следующий
Общество эндокринологов BES 2013Общество эндокринологов
Просмотреть другие тома
Резюме Абстрактная книга Программа Редакторы томов эпипостеры Тезисы
Прикрепите ePoster к своему тезису
| Отказ от ответственности
Нет недавних поисков.
Эндокринные рефераты Google ученый Паб Мед
Эндокринные рефераты Google ученый Паб Мед
Эндокринные рефераты Google ученый Паб Мед
Эндокринные рефераты Google ученый Паб Мед
Эндокринные рефераты Google ученый Паб Мед
Эндокринные рефераты Google ученый Паб Мед
Ингибирование экспрессии фосфопротеина р18 антисмысловой РНК отменяет трансформированный фенотип лейкемических клеток1 | Исследование рака
Пропустить пункт назначения Nav
Статьи|
01 марта 1996 г.
Сима Джеха;
Сян-Нонг Луо;
Милослав Беран;
Акоп Кантарджян;
Джордж Ф. Атве
Информация об авторе и статье
2
Кому следует направлять запросы на перепечатку, в отделение гематологии, ящик 1079, Медицинская школа Маунт-Синай, One Gustave L. Levy Place, New York, NY 10029. Телефон: (212) 241- 5293; Факс: (212) 369-8375.
Полученный: 11 сентября 1995 г.
Принято: 15 января 1996
Номер в сети: 1538-7445
Номер для печати: 0008-5472
©1996 Американская ассоциация исследований рака.
1996
Американская ассоциация исследований рака, Inc.
Cancer Res (1996) 56 (6): 1445–1450.
История статьи
Получено:
11 сентября 1995 г.
Принято:
15 января 1996 г.
Цитирование
Сима Джеха, Сян-Нонг Луо, Милослав Беран, Агоп Кантарджян, Джордж Ф.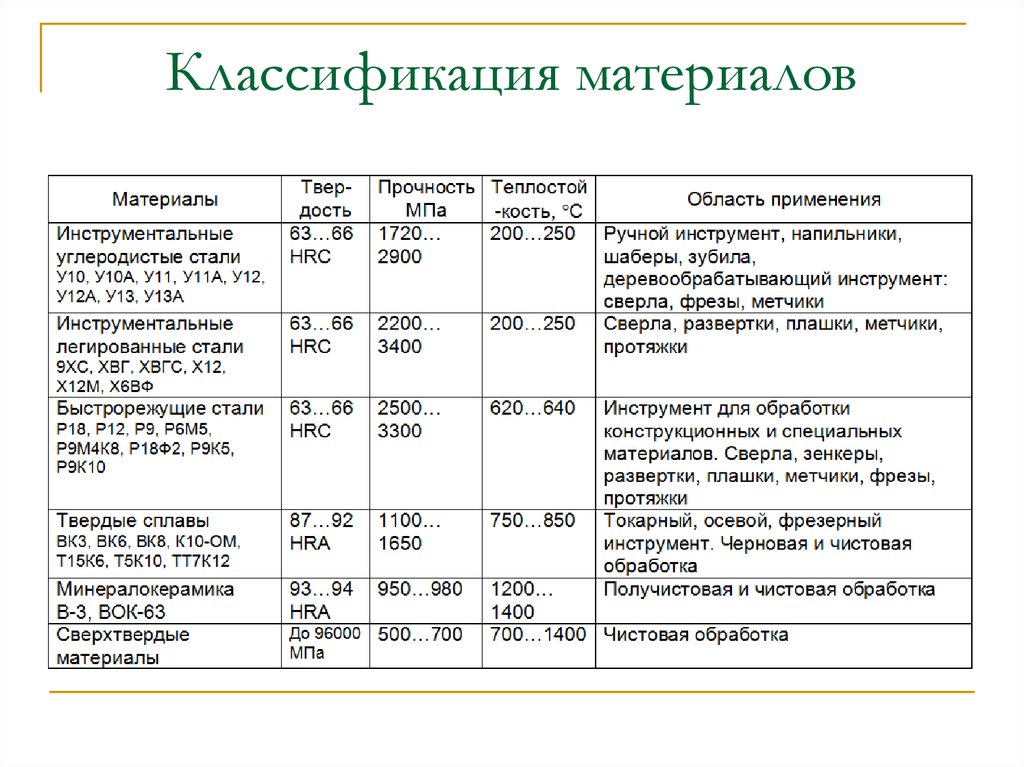 Атве; Ингибирование экспрессии фосфопротеина p18 антисмысловой РНК устраняет трансформированный фенотип лейкемических клеток 1 . Cancer Res 15 марта 1996 г.; 56 (6): 1445–1450.
Атве; Ингибирование экспрессии фосфопротеина p18 антисмысловой РНК устраняет трансформированный фенотип лейкемических клеток 1 . Cancer Res 15 марта 1996 г.; 56 (6): 1445–1450.
Скачать файл цитаты:
Расширенный поиск
Фосфопротеин р18 первоначально был идентифицирован на основании его очень высокого уровня экспрессии в лейкемических клетках различных линий. Изменения уровня накопления и фосфорилирования p18, связанные с индукцией дифференцировки лейкемических клеток, предполагают потенциальную роль этого фосфопротеина в клеточной пролиферации и дифференцировке и, возможно, в злокачественной трансформации. Недавние исследования показали, что p18 играет важную роль в развитии клеточного цикла, выступая в качестве субстрата для p34 9.0005 киназа cdc2 . Эти исследования показали, что ингибирование экспрессии p18 в лейкемических клетках приводит к замедлению роста и накоплению клеток в G 2 -M. В этом исследовании мы изучаем потенциальную роль p18 в клеточной трансформации, исследуя влияние ингибирования экспрессии p18 на злокачественный фенотип клеток эритролейкемии K562. Эти исследования показывают, что антисмысловое ингибирование экспрессии p18 в лейкемических клетках приводит к остановке роста при более низкой плотности насыщения, потере независимости от сыворотки и потере независимого от прикрепления роста in vitro . Кроме того, ингибирование экспрессии p18 приводит к заметному ингибированию туморогенности лейкозных клеток in vivo в мышиной модели с тяжелым комбинированным иммунодефицитом. Эти исследования демонстрируют, что высокий уровень экспрессии р18 в лейкемических клетках необходим для поддержания трансформированного фенотипа, и позволяют предположить, что р18 является потенциальной мишенью для противолейкозных вмешательств.
Недавние исследования показали, что p18 играет важную роль в развитии клеточного цикла, выступая в качестве субстрата для p34 9.0005 киназа cdc2 . Эти исследования показали, что ингибирование экспрессии p18 в лейкемических клетках приводит к замедлению роста и накоплению клеток в G 2 -M. В этом исследовании мы изучаем потенциальную роль p18 в клеточной трансформации, исследуя влияние ингибирования экспрессии p18 на злокачественный фенотип клеток эритролейкемии K562. Эти исследования показывают, что антисмысловое ингибирование экспрессии p18 в лейкемических клетках приводит к остановке роста при более низкой плотности насыщения, потере независимости от сыворотки и потере независимого от прикрепления роста in vitro . Кроме того, ингибирование экспрессии p18 приводит к заметному ингибированию туморогенности лейкозных клеток in vivo в мышиной модели с тяжелым комбинированным иммунодефицитом. Эти исследования демонстрируют, что высокий уровень экспрессии р18 в лейкемических клетках необходим для поддержания трансформированного фенотипа, и позволяют предположить, что р18 является потенциальной мишенью для противолейкозных вмешательств.
1
Работа выполнена при поддержке гранта NIH PHS HL-42919 (GFA) и грант Центра общих клинических исследований NIH M01-RR00071 (Медицинская школа горы Синай).
Этот контент доступен только в формате PDF.
Пептид P18, функциональный фрагмент фактора пигментного эпителия, ингибирует ангиогенез при гепатоцеллюлярной карциноме посредством модулирования сигнального пути VEGF/VEGFR2 ингибитор. Это исследование было направлено на определение антиангиогенной биологической активности пептида Р18 при гепатоцеллюлярной карциноме (ГЦК) и выяснение основного механизма. Анализы роста опухоли ксенотрансплантата продемонстрировали, что пептид P18 подавляет ангиогенез ГЦК in vivo. Анализы заживления ран, Transwell и Matrigel-культуры показали, что пептид P18 ингибирует миграцию клеток и образование трубок эндотелиальных клеток (ЭК) in vitro. Жизнеспособность клеток и апоптоз, оцененные с помощью набора Cell Counting Kit-8 (CCK-8) и анализов проточной цитометрии, позволили предположить, что пептид P18 ингибирует ангиогенез, индуцируя апоптоз EC.
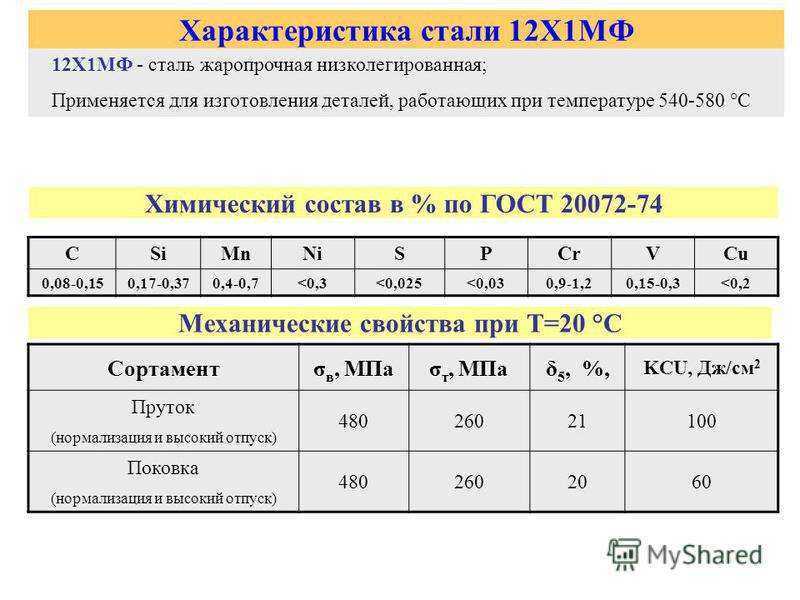 Молекулы, связанные с ангиогенезом и сигнальной трансдукцией, проанализированные с помощью вестерн-блоттинга, показали, что пептид P18 нацелен на рецептор 2 фактора роста эндотелиальных клеток сосудов (VEGFR2) на ЭК. В заключение, ингибируя фосфорилирование VEGFR2, пептид P18 модулирует передачу сигналов между VEGF/VEGFR2 и подавляет активацию каскадов PI3K/Akt, что приводит к усилению митохондриально-опосредованного апоптоза и антиангиогенной активности. Эта биоактивность пептида Р18 может представлять собой новую терапевтическую стратегию лечения ГЦК.
Молекулы, связанные с ангиогенезом и сигнальной трансдукцией, проанализированные с помощью вестерн-блоттинга, показали, что пептид P18 нацелен на рецептор 2 фактора роста эндотелиальных клеток сосудов (VEGFR2) на ЭК. В заключение, ингибируя фосфорилирование VEGFR2, пептид P18 модулирует передачу сигналов между VEGF/VEGFR2 и подавляет активацию каскадов PI3K/Akt, что приводит к усилению митохондриально-опосредованного апоптоза и антиангиогенной активности. Эта биоактивность пептида Р18 может представлять собой новую терапевтическую стратегию лечения ГЦК.Введение
Ангиогенез относится к образованию новой крови
сосуды из существующих капилляров или путем внутривенной разработки.
Полный процесс ангиогенеза включает множество клеток
и молекулы, которые играют роль в базальной мембране сосудов
деградации, а также в последующем эндотелиальных клетках сосудов
активация, пролиферация, миграция и реконструкция (1). В процессе развития опухоли
ангиогенез является необходимой предпосылкой и регулируется
баланс между проангиогенными факторами, такими как сосудистая
фактор роста эндотелиальных клеток (VEGF), основной рост фибробластов
фактор (bFGF) и ангиопоэтин, а также антиангиогенные факторы, такие как
как тумстатин, эндостатин и тромбоспондин-1 (2–4). Гепатоцеллюлярная карцинома (ГЦК) является одним из наиболее распространенных видов
карцинома и характеризуется обогащенным кровоснабжением.
прогрессирование ангиогенеза имеет жизненно важное значение для возникновения опухоли и
развития и тесно связан с метастазированием ГЦК, инвазией
и резистентность к терапии (5,6). Таким образом,
несколько антиангиогенных препаратов (например, сорафениб)
рекомендуется для клинического применения в качестве вариантов лечения
пациенты с поздней стадией ГЦК (7,8).
Гепатоцеллюлярная карцинома (ГЦК) является одним из наиболее распространенных видов
карцинома и характеризуется обогащенным кровоснабжением.
прогрессирование ангиогенеза имеет жизненно важное значение для возникновения опухоли и
развития и тесно связан с метастазированием ГЦК, инвазией
и резистентность к терапии (5,6). Таким образом,
несколько антиангиогенных препаратов (например, сорафениб)
рекомендуется для клинического применения в качестве вариантов лечения
пациенты с поздней стадией ГЦК (7,8).
Фактор пигментного эпителия (PEDF) представляет собой
многофункциональный гликопротеин, относящийся к семейству
неингибирующие серпины. Среди природных антиангиогенных
факторы, PEDF считается эффективным ингибитором ангиогенеза
(9,10). Однако PEDF состоит из нескольких
функциональные фрагменты, отвечающие за несколько функций,
такие как ингибирование ангиогенеза и повышение выживаемости и
нейродифференциация. Предыдущие исследования показали, что
антиангиогенный функциональный фрагмент PEDF расположен в
Nh3-концевой поверхностный эпитоп (34-мерные аминокислотные остатки 24–57,
PEDF-34mer), в то время как средства, способствующие выживанию и нейродифференциации
функциональные фрагменты расположены на соседнем эпитопе (44-мерный
остатки 58–101, 44-мер PEDF) (11–13). Исследования показали, что PEDF ингибирует развитие нескольких
злокачественные опухоли, такие как карцинома легкого, остеосаркома и
карцинома поджелудочной железы (14–16). Однако, хотя в естественных условиях
сверхэкспрессия полноразмерного PEDF полезна для ингибирования
рост опухоли, его применение в клинической практике ограничено.
из-за его низкой стабильности и иммуногенности. Поэтому коротко
но стабильные пептиды, полученные из PEDF, которые столь же эффективны при
ингибирование ангиогенеза, так как полноразмерные фрагменты PEDF могут
имеют большее практическое значение в клинической практике. Пептид Р18 представляет собой
ангиоингибиторный эпитоп 34-мерного PEDF (18-мерные остатки 40–57)
который был предложен в качестве биоактивного антиангиогенного фрагмента.
Предыдущее исследование показало, что пептид P18 проявлял активность
аналогично полному PEDF в предстательной железе и почках
рак (17). Тем не менее
влияние пептида Р18 на ангиогенез при ГЦК и его
применимость в терапии опухолей остаются неясными.
Исследования показали, что PEDF ингибирует развитие нескольких
злокачественные опухоли, такие как карцинома легкого, остеосаркома и
карцинома поджелудочной железы (14–16). Однако, хотя в естественных условиях
сверхэкспрессия полноразмерного PEDF полезна для ингибирования
рост опухоли, его применение в клинической практике ограничено.
из-за его низкой стабильности и иммуногенности. Поэтому коротко
но стабильные пептиды, полученные из PEDF, которые столь же эффективны при
ингибирование ангиогенеза, так как полноразмерные фрагменты PEDF могут
имеют большее практическое значение в клинической практике. Пептид Р18 представляет собой
ангиоингибиторный эпитоп 34-мерного PEDF (18-мерные остатки 40–57)
который был предложен в качестве биоактивного антиангиогенного фрагмента.
Предыдущее исследование показало, что пептид P18 проявлял активность
аналогично полному PEDF в предстательной железе и почках
рак (17). Тем не менее
влияние пептида Р18 на ангиогенез при ГЦК и его
применимость в терапии опухолей остаются неясными.
VEGF является сильнодействующим проангиогенным цитокином,
Также известно, что он способствует пролиферации и выживанию эндотелиальных
клетки (ЭК), а также проницаемость сосудов (18,19).
VEGF экспрессируется в васкуляризированных тканях и имеет решающее значение для
нормальный и патологический ангиогенез. Веские доказательства
участие VEGF в индукции опухолевой пролиферации,
метастазирование и ангиогенез (3,20,21).
VEGF165, преобладающая изоформа VEGF у людей, передает сигналы через
три рецептора: fms-подобная тирозинкиназа (flt-1, также VEGFR1), KDR
продукт гена (KDR, также VEGFR2) и продукт гена flt4 (flt-4,
также VEGFR3). Среди этих трех рецепторов VEGFR2 является основным.
рецептор для индуцированной VEGF передачи сигналов в эндотелиальных клетках (3, 22–24).
Предыдущие исследования выявили VEGFR2 не только в сосудистых
эндотелиальных клеток, но и его гиперэкспрессию во многих типах
злокачественные солидные опухоли (25). На
связываясь с VEGF, VEGFR2 подвергается аутофосфорилированию и становится
активирован. Фосфорилирование Tyr1175 позволяет связываться с p85.
субъединица киназы PI-3 (PI3K), что приводит к активации
Сигнальный путь PI3K/Akt (26).
Эта сигнализация VEGFR2 необходима для выполнения
Стимулируемая VEGF пролиферация, хемотаксис и прорастание, а также
для выживания культивируемых эндотелиальных клеток in vitro и
ангиогенез in vivo (20,24).
Имеющиеся данные указывают на то, что VEGFR2 является
цель PEDF (9,27). Однако может ли пептид Р18
блокировать передачу сигнала VEGF/VEGFR2 и в конечном итоге привести к
ингибирование ангиогенеза при ГЦК остается неясным. Таким образом, мы
разработали настоящее исследование, чтобы определить механизм, с помощью которого
Пептид Р18 ингибирует ангиогенез при ГЦК. Мы заметили, что P18
пептид противодействовал биоактивности VEGF и подавлял клеточную
активности за счет снижения секреции кадгеринов (VE-кадгерин и
Е-кадгерин) и матриксные металлопротеиназы (матриксные
металлопротеиназа-2, ММП-2 и ММП-9) в обеих пупочных венах человека
эндотелиальные клетки микрососудов (HUVEC) и гепатома HepG2
клеток in vitro, что привело к подавлению
инвазивность и проангиогенез эндотелиальных клеток. Более того,
наша модель опухоли ксенотрансплантата также предоставила доказательства того, что P18
пептид подавляет фосфорилирование VEGFR2 и ингибирует
ангиогенез ГЦК in vivo. Мы продемонстрировали, что P18
пептид может действовать на ЭК, модулируя передачу сигналов VEGF/VEGFR2.
пути и индуцирует каскады PI3K/Akt, что приводит к
апоптоз ЭК и снижение неоваскуляризации.
Материалы и методы
Клеточные линии и культуры
Пупочная вена человека и эндотелий микрососудов
клетки (HUVEC) и клетки HepG2 линии клеток HCC человека были приобретены
из Центра заповедника типичной культуры Китая (Шанхай, Китай)
и культивировали при 5% CO2 в среде ЕСМ-2 (ScienCell
Research Laboratories, Карлсбад, Калифорния, США) с добавлением 5%
эмбриональная бычья сыворотка (FBS, Gibco, Grand Island, NY, USA), 100 ЕД/мл
пенициллин, 100 мкг/мл стрептомицина и 1% рост эндотелиальных клеток
добавка (ECGS, ScienCell Research Laboratories). ГЦК человека
клеточная линия HepG2 была получена из Американской Типовой Культуры
Коллекция (Роквилл, Мэриленд, США) и культивируется в модифицированной Дульбекко культуре. среда Игла (DMEM, Hyclone, Thermo Fisher Scientific Inc.,
Логан, Юта, США) с добавлением 10% FBS (Gibco), 100 ЕД/мл
пенициллин и стрептомицин 100 мкг/мл. Использовали гипоксический инкубатор.
для имитации гипоксических условий (1% O2, 5% CO2
и 94% N2).
Антитела и реагенты
Антитела против VEGFR2, фосфорилированные (p)-VEGFR2
(Tyr1175), PI3K, p-PI3K p85 (Tyr458)/p55 (Tyr199), Akt, p-Akt
(Ser473), Bax, Bcl-2, каспаза-3, расщепленная каспаза-3, ММП-2, ММП-9
и CD31 были приобретены у Cell Signaling Technology (Danvers,
Массачусетс, США). Антитела против VEGF, VE-кадгерина, Е-кадгерина, Ki67,
GAPDH и β-актин были приобретены у Abcam (Кембридж, Великобритания).
Конъюгированные с пероксидазой хрена (HRP) козьи антикроличьи IgG и
Конъюгированные с HRP мышиные IgG были приобретены в Beyotime Institute of
Биотехнология (Цзянсу, Китай). Рекомбинантный пептид Р18 (>95%
чистоты) был приобретен у GL Biochem Ltd. (Шанхай, Китай) и
характеризуется масс-спектрометрией. Пептиды ацетилировали по
их концы Nh3 и амидированы на их концах COOH для стабильности. SU1498, селективный ингибитор VEGFR2, был приобретен у Abcam.
Рекомбинантный человеческий VEGF (VEGF165) и рекомбинантный человеческий IGF-1 были
получен от PeproTech (Роки Хилл, Нью-Джерси, США). Матригель был
приобретен у BD Biosciences (Сан-Хосе, Калифорния, США).
Анализ жизнеспособности клеток
HUVEC (5×103/лунка) или клетки HepG2
(5×103/лунку) высевали в три 96-луночные планшеты. После
прикрепления клеток супернатанты в чашках заменяли на
ЕСМ, содержащий градиент концентрации пептида Р18 (10, 20,
40, 80, 160, 320, 640 и 1280 нМ). Таблички были пронумерованы как
планшет 1, планшет 2 и планшет 3 и инкубировали при 37°C при 5%
СО2. Жизнеспособные клетки определяли количественно с помощью набора Cell Counting Kit-8.
(CCK-8, Dojindo Molecular Technologies, Кумамото, Япония)
различные моменты времени (таблица 1:24 ч, пластина 24:8 ч и пластина 3:72 ч).
Оптическую плотность (ОП) при 450 нм измеряли с помощью спектрометра Spectra Max.
190 (Molecular Devices, Саннивейл, Калифорния, США).
Вестерн-блот анализ
Клетки или образцы замороженных опухолей лизировали в
буфер для холодного лизиса RIPA (Beyotime, Пекин, Китай) с 1 нМ
фенилметилсульфонилфторид. Набор для анализа белков BCA (Beyotime)
использовали для измерения концентрации белковых образцов. Общий
белок (20–25 мкг) разделяли с помощью гелей SDS-PAGE и переносили
к мембранам PVDF (Millipore, Billerica, MA, USA). Мембраны
блокировали 5% обезжиренным молоком на 2 ч при комнатной температуре и
затем инкубировали с первичными антителами при 4°С в течение ночи согласно
к инструкциям производителя. Затем были мембраны
инкубируют с конъюгированными с пероксидазой хрена вторичными антимышиными или кроличьими IgG
антитела (Бейотим) в течение 2 часов при комнатной температуре, а затем
обнаружение с помощью иммуноблотинга с усиленной хемилюминесценцией (ECL)
реагенты для обнаружения (Millipore). Интенсивность белковых полос была
количественно определено с помощью денситометрического анализа с использованием программного обеспечения ImageJ
(Национальные институты здоровья, Бетесда, Мэриленд, США).
Анализ роста опухоли ксенотрансплантата
Клетки HepG2 (5×106/0,1 мл) в PBS инокулировали в область спины возле передней ноги 4-недельного BALB/c голые мыши (Shanghai Laboratory Animal Company, Шанхай, Китай). За мышами наблюдали до тех пор, пока опухоль не достигала объема 100 мм3. Затем мышей рандомизировали на три группы (5 в каждой группе) и отмечены. Две экспериментальные группы были внутрибрюшинно вводили пептид Р18 (растворенный с 0,9% физиологическим раствором) в дозах 0,1 мг/кг (доза/тело мыши). вес) или 0,5 мг/кг. Контрольную группу лечили тем же объем 0,9% физиологический раствор (0,9% NS). Наблюдали за ростом опухоли по длине и ширине опухоли, полученной из наружного замеры каждые 2 дня. Объем опухоли определяли по следующему уравнению: объем (мм3) = (длина × ширина3)/2. Через четырнадцать дней после первой инъекции мышей умерщвляли, а опухоли вырезали и взвешивали. Все эксперименты были проведены после одобрения этики комитет больницы Цяньфошань.
Иммунофлуоресцентный анализ (ИФ)
Ксеногенные опухоли были заморожены в жидкости
азота сразу после умерщвления и последующего замораживания мышей. на срезах толщиной 5 мкм. Срезы фиксировали в 4% параформальдегиде.
в течение 20 мин. После блокировки в 5% БСА в течение 1 ч срезы
инкубировали при 4°C в течение ночи с козьим поликлональным анти-VEGF (Novus,
Сан-Диего, США) и моноклональный антифосфорилированный VEGFR2 кролика.
Затем клетки или мышиные срезы инкубировали с ослиными антикозьими антителами.
С FITC-меченым (Abcam) и осла против кролика с TRITC-меченым (Abcam)
вторичные антитела в течение 2 ч и окрашивали DAPI (Abcam).
Флуоресцентные изображения тканей фотографировали с помощью светового
микроскоп, и двойное иммунофлуоресцентное окрашивание объединяли с помощью
Программное обеспечение Image-Pro Plus.
Иммуногистохимический анализ (ИГХ)
Простые опухоли фиксировали в 10% формалине, заливали
в парафине, а затем подвергают иммуногистохимии.
срезы депарафинизировали в ксилоле градуированной концентрации и регидратировали в
этанолом степени очистки, а затем 3 раза промывали PBS. После
индуцированный нагреванием поиск антигена в цитратном буфере, эндогенный
Пероксидаза ингибируется обработкой 3% перекисью водорода при
комнатной температуре в течение 10 мин с последующим промыванием 3 раза PBS. Затем срезы инкубировали с первичным анти-VE кадгерином,
анти-CD31 и анти-Ki67 антитела в течение ночи. После мытья с
PBS, срезы обрабатывали пероксидазой хрена.
(HRP)-конъюгированный козий антикроличий IgG в течение 1 ч при 37°C. Отрицательный
контрольные срезы инкубировали с PBS вместо первичных
антитело.
Анализ заживления ран
HUVEC высевали в 6-луночные планшеты. Когда клетки слились на 90%, линию раны сделали с помощью 10-мкл пластиковой наконечник пипетки. Затем клетки инкубировали в бессывороточном ЕСМ-2. среда с VEGF (8 нг/мл) с пептидом Р18 или без него (0,2 мкМ). Раневые процессы фотографировали в моменты времени 0, 12 и 24 ч, а расстояние миграции клеток определяли количественно с помощью анализ изображений.
Анализ миграции и инвазии
Система миграции клеток Transwell (8 мкм, Corning
Inc., Корнинг, Массачусетс, США), покрытые матригелем, использовали для выполнения
анализ миграции и инвазии клеток. HUVEC
(1×105/лунку) или клетки HepG2 (5×104/лунку)
добавляют в верхнюю камеру планшета Transwell и 500 мкл
бессывороточная среда с VEGF или без него (8 нг/мл) и P18
пептид (0,2 мкМ) добавляли в нижнюю камеру. Через 48 часов
инкубации мигрировавшие клетки фиксировали 95% метанол и
окраска 0,1% кристаллическим фиолетовым в течение 30 мин с последующей отмывкой 5
раз с PBS.
Анализ формирования пробирок
Каждую лунку 96-луночного планшета покрывали 50 мкл Матригель. HUVEC (1×104/лунку) высевали в лунки. после затвердевания матригеля. Клетки в экспериментальные группы обрабатывали пептидом Р18 (0,2 мкМ), а контрольные группы обрабатывали равным объемом PBS. Клетки инкубировали при 37°C с 5% CO2 в течение 6 ч, а затем сфотографировано с помощью светового микроскопа (Olympus, Токио, Япония).
Анализ клеточного апоптоза
HUVEC (4×105/лунка) или клетки HepG2
(4×105/лунка) обрабатывали с P18 или без него.
пептида и инкубируют в течение 24 часов. Затем культивированные клетки
суспендировали в PBS, подсчитывали и ресуспендировали в буфере для связывания.
Аннексин V-FITC и йодид пропидия (PI; NeoBioscience Ltd.,
Шэньчжэнь, Китай) использовали для определения скорости апоптоза
клеток с помощью проточной цитометрии.
Статистический анализ
Данные были проанализированы с помощью программного обеспечения SPSS (версия 17.0, SPSS China, Шанхай, Китай) и выражается как среднее значение ± стандартное отклонение (SD). Для сравнения между 2 группами и однофакторный дисперсионный анализ. используется для сравнения нескольких групп. P-значения <0,05 считались статистически значимыми результатами.
Результаты
Пептид Р18 ингибирует пролиферацию ЭК in vitro
Пептид Р18 был синтезирован в соответствии с аминокислотная последовательность, показанная на фиг. 1А. Анализ доза-реакция, проведенный с помощью анализа CCK-8, подтвердил IC50 ~320 нмоль/л для пептида Р18 in vitro (рис. 1Б). В качестве биомаркера для уровень апоптоза, Bcl-2 и Bax были обнаружены западными промокание. Мы обрабатывали HUVEC различными концентрациями P18. пептида и наблюдал дозозависимое подавление Bcl-2, но активация Bax (рис. 1C).
Рисунок 1. Пептид P18 ингибирует EC
размножение in vitro. |
Пептид Р18 подавляет рост опухоли и ангиогенез ГЦК in vivo
Для оценки влияния пептида Р18 на опухоль
рост in vivo, мы разработали анализ роста опухоли ксенотрансплантата
с клетками HepG2 у голых мышей. Две экспериментальные группы получили
Пептид Р18 в дозе 0,1 мг/кг или 0,5 мг/кг, в то время как контроль
группа получала тот же объем 0,9% физиологического раствора (0,9% NS).
средний объем опухоли составил 47,40% у получавших P18 (0,1 мг/кг)
группе и 20,64% в группе, получавшей P18 (0,5 мг/кг), нормализованной до
контрольная группа (рис. 2А и В).
В качестве отличительных признаков ангиогенеза уровни экспрессии CD31 и
VE-кадгерин в опухолевых тканях выявляли с помощью ИГХ-анализа, который
выявили постепенное снижение CD31 и VE-кадгерина с увеличением
дозировка пептида Р18 (рис. 2C-E).
Кроме того, вестерн-блоттинг показал значительное
снижение уровней экспрессии CD31 и VE-кадгерина
(рис. 2F и G).
Рисунок 2. Пептид P18 подавляет опухоль
рост и ангиогенез ГЦК in vivo. (A) Опухолевая ткань на
14-й день после введения 0,9% НС или различных доз Р18. (Б)
Кривая подавления роста опухоли представлена в виде объема опухоли после
лечение 0,9% NS или различными дозами P18 (среднее значение ± стандартное отклонение). (С)
Результаты окрашивания ИГХ на CD31 и VE-кадгерин в опухолевых тканях
(увеличение, ×200). (D) Данные представляют МВД в опухолевых тканях.
(среднее значение ± стандартное отклонение, *P<0,05, ***P<0,01). МВД определяли с помощью ИГХ
окрашивание эндотелиально-специфическим антителом против CD31
(увеличение, ×200). |
Пептид Р18 ингибирует фосфорилирование VEGFR2 in vivo
Уровни экспрессии VEGF и p-VEGFR2 в опухоли ткани были обнаружены с помощью IF-анализа. Результаты показали, что наблюдалось дозозависимое подавление экспрессии VEGF с последующим снижением уровня фосфорилирования VEGFR2 в Группы, получавшие Р18 (рис. 3А). Вестерн-блоттинг также показал, что пептид Р18 снижает экспрессию VEGF и подавляет фосфорилирование VEGFR2 в опухолевых тканях HCC (фиг. 3B и C).
Рисунок 3.Пептид P18 ингибирует фосфорилирование VEGFR2 in vivo. (А) Выражение и распределения VEGF и p-VEGFR2 в тканях ксенотрансплантата опухоли. определяется с помощью иммунофлуоресцентной микрофотографии (увеличение, ×200). (B) Уровни экспрессии VEGFR2, p-VEGFR2 и VEGF в опухоли ткани были обнаружены вестерн-блоттингом. (С) Плотность полосы в вестерн-блоттинге количественно определяли и нормализовали к этому GAPDH (среднее значение ± стандартное отклонение). |
Пептид P18 подавляет жизнеспособность ЭК путем противодействие биологической активности VEGF in vitro
Анализы заживления ран и анализы Transwell были
для изучения влияния пептида Р18 на
миграционная и инвазивная способность ЭК в присутствии VEGF, а также
результаты показали, что пептид P18 значительно ингибирует
Миграция и инвазия HUVEC при концентрации 0,2 мкМ (рис. 4A-D). Результаты вестерн-блоттинга
предположили, что пептид P18 достиг этой биологической активности в первую очередь
за счет снижения уровней экспрессии MMP-2 и MMP-9в
HUVEC (рис. 4E и F). Для дальнейшего
исследовать антиангиогенную активность пептида Р18 в
vitro мы высевали HUVEC на 96-луночные планшеты, покрытые матригелем, в течение 8
ч с использованием дополнительного VEGF (8 нг/мл) в качестве положительного контроля.
результаты показали, что VEGF может усиливать ангиогенез HUVEC.
но пептид Р18 обратил этот эффект и ингибировал ангиогенез.
in vitro (рис. 5А и В). В
в то же время вестерн-блоттинг подтвердил, что дополнительный VEGF
может повышать уровень экспрессии VE-кадгерина и E-кадгерина в
HUVEC, в то время как пептид P18 противодействовал этой биологической активности.
(Рис. 5C и D).
Рисунок 4. Пептид P18 подавляет клетки
жизнеспособность HUVEC и противодействует биологической активности VEGF в
витро. (A) Влияние пептида P18 на миграцию HUVEC
были проанализированы методом заживления ран. (B) Данные представляют
расстояние миграции в процентах от контрольной группы в ране
анализ заживления (среднее значение ± стандартное отклонение, *P<0,05, ***P<0,01). |
Рисунок 5. Пептид P18 ингибирует ангиогенез
способность HUVEC путем противодействия биологической активности VEGF в
витро. (A) HUVEC культивировали с VEGF или без него, а
Пептид Р18 (0,2 нМ). Анализировали ангиогенную способность HUVEC.
по анализу трубообразования. (B) Данные представляют количество закрытых
пробирки в контрольной и экспериментальной группах (среднее значение ± стандартное отклонение,
*Р<0,05, ***Р<0,01). |
Пептид Р18 индуцирует апоптоз ЭК in vitro
Поскольку данное исследование предварительно определило IC50 ~320 нмоль/л для пептида Р18 in vitro, далее мы культивировали HUVEC в среде DMEM с VEGF или без него. Через 12 часов к раствору добавляли растворенный в PBS пептид P18 (0,32 мкМ). опытной группе, и клетки культивировали еще 24 часа. Аннексин V-FITC/PI использовали проточную цитометрию и вестерн-блоттинг. для определения уровней апоптоза HUVEC. Результаты, достижения постоянно указывалось, что дополнительное количество VEGF может подавлять апоптоз HUVEC, и этот процесс может быть обращен вспять с помощью P18. пептид (фиг. 6A-D).
Рисунок 6.Пептид Р18 индуцирует апоптоз HUVEC in vitro. (A) HUVEC культивировали с или без VEGF и пептид Р18 (0,32 нМ) в течение 24 часов. Уровни апоптоза были оценивают с помощью проточной цитометрии. (B) Данные представляют процент клеток с ранним и поздним апоптозом и выражается как среднее ± SD, *P<0,05, ***P<0,01. (C) Уровни экспрессии Bcl-2 и Bax в HUVEC анализировали вестерн-блоттингом после культура с VEGF или без него (8 нг/мл) и пептидом Р18 (0,32 нМ) в течение 24 часов. (D) Плотность каждой полосы в вестерн-блоттинге был количественно определен и нормализован по отношению к GAPDH (среднее значение ± стандартное отклонение, *Р<0,05, ***Р<0,01). |
Пептид P18 подавляет клетки HepG2 жизнеспособность за счет подавления миграции клеток, а не ингибирования клеток пролиферация, вызывающая апоптоз
Для изучения
влияние пептида P18 на миграционную способность клеток HepG2
в присутствии VEGF. Результаты показали, что пептид P18
ингибирует миграцию опухолевых клеток и противодействует
биоактивность VEGF in vitro (фиг. 7А и В). Уровни экспрессии MMP-2,
ММП-9и E-кадгерин в клетках HepG2, обработанных VEGF (8 нг/мл) и
пептид Р18 (0,32 мкМ) определяли вестерн-блоттингом.
Высокий уровень экспрессии MMP-2 и MMP-9, стимулируемый VEGF в
Клетки HepG2 подавлялись пептидом P18, в то время как клетки
экспрессия E-кадгерина была слегка увеличена в P18
группы, обработанные пептидами, которые способствуют подавлению клеточного
миграции (рис. 7C и D). Однако,
после обработки различными концентрациями пептида Р18
в разное время существенных различий не наблюдалось.
пролиферацию клеток HepG2 в анализах CCK-8 (фиг. 7E). Кроме того, результаты
Аннексин V-FITC/PI проточная цитометрия и вестерн-блоттинг не показали
значительная разница в апоптозе клеток HepG2 между контролем
и группы, получавшие пептид P18 (фиг. 7F).
и Г). Следует отметить, что эксперименты на рис. 7E-G проводились в отсутствие
Клетки VEGF, но HepG2 культивировали в среде DMEM с 10% FBS.
Поскольку клетки HepG2 могут поддерживать высокий уровень пролиферации при
это состояние без необходимости дополнительного введения VEGF, мы считаем
что клеток в нормальной сывороточной среде достаточно в качестве контрольной группы
(28,29). Приведенные выше результаты свидетельствуют о том, что
Пептид P18 подавляет жизнеспособность клеток HepG2 путем подавления клеточного
миграции, а не за счет ингибирования пролиферации клеток и индукции
апоптоз.
Рисунок 7. Пептид P18 подавляет клетки HepG2
жизнеспособность за счет подавления миграции клеток, а не ингибирования клеток
пролиферации и индукции апоптоза. (А) Пептид Р18
противодействует биологической активности VEGF и ингибирует миграцию
Клетки HepG2, оцененные с помощью анализов Transwell. (B) Данные представляют
количество клеточных метастазов (среднее значение ± стандартное отклонение, *P<0,05, ***P<0,01).
(C) Пептид P18 подавлял уровни экспрессии MMP-2 и
ММП-9в клетках HepG2 по оценке вестерн-блоттинга. (Г)
плотность каждой полосы в вестерн-блоттинге была определена количественно и
нормализовано по отношению к GAPDH (среднее значение ± стандартное отклонение, * P <0, 05, *** P <0, 01).
(E) Титрование дозы пролиферации клеток HepG2. |
Пептид Р18 индуцирует апоптоз ЭК посредством блокирует VEGF/VEGFR2 и индуцирует передачу сигналов PI3K/Akt путь
SU1498 является селективным ингибитором VEGFR2, который
главный рецептор VEGF-индуцированной биологической активности в эндотелии
клетки (23,30). В этом исследовании мы культивировали HUVEC в
бессывороточной среде с дополнительным VEGF и использовали SU1498 в качестве
положительный контроль на пептид Р18. HUVEC, культивированные в среде
с 10% FBS или VEGF служили в качестве отрицательного контроля. Вестерн-блот
результаты показали, что пептид Р18 ингибирует VEGF-индуцированный
фосфорилирование VEGFR2 аналогично фосфорилированию SU1498
(рис. 8А). Для дальнейшей демонстрации
механизм, с помощью которого P18 ингибирует ангиогенез, мы лечили HUVECs
в бессывороточной среде в течение 24 ч, а затем заменили среду на
дополнительный VEGF с пептидом P18 или без него (0,2 мкМ). ИФР-1,
активатор пути PI3K/Akt, использовали в качестве отрицательного контроля
(31). Как нисходящая сигнализация
ось VEGFR2, активация PI3K и Akt через
фосфорилирование в HUVEC значительно подавлялось после
обработка пептидом Р18 в течение 90 мин, которую можно было обратить
с присутствием IGF-1 (0,2 нг/мл) (фиг. 8B и C). После культивирования с
Пептид Р18 или ИФР-1 в течение 4 ч, Bcl-2/Bax в качестве биомаркеров для
степень апоптоза и их последующие молекулы-мишени, расщепленные
каспаза-3/каспаза-3 также были обнаружены вестерн-блоттингом. результаты показали, что пептид P18 подавлял экспрессию
Bcl-2 индуцировал экспрессию Bax и усиливал протеолитическую
процессинг неактивной каспазы-3 в расщепленную каспазу-3. Однако,
этим эффектам противодействовал IGF-1 (фиг. 8D и E). Эти результаты показали
что деактивация сигнального пути VEGF/VEGFR2 и его
индуцированные каскады PI3K/Akt могут быть защитным механизмом, участвующим в
индуцированный пептидом Р18 апоптоз HUVEC и его антиангиогенный
Мероприятия.
Рисунок 8. Пептид P18 блокирует VEGF/VEGFR2
сигнальный путь и индуцирует каскад PI3K/Akt. (А) стр.18
пептид ингибировал фосфорилирование VEGFR2 аналогично
к SU1498. Данные представляют относительную плотность полос и процент
p-VEGFR2/VEGFR2, выраженное как среднее значение ± стандартное отклонение. (В и С) P18
пептид ингибировал индуцированное VEGF фосфорилирование PI3K и Akt,
который может быть реверсирован ИФР-1. Данные представляют относительную полосу
плотности и нормированы на GAPDH (среднее ± стандартное отклонение, ***P<0,01). |
Обсуждение
Как тип богато васкуляризированной солидной опухоли,
возникновение и развитие ГЦК во многом зависит от
ангиогенез (5). Следовательно,
антиангиогенное лечение стало клинически значимым
терапевтические варианты лечения пациентов с ГЦК (7). Ангиогенез в первую очередь зависит от
проангиогенные и антиангиогенные факторы. PEDF, тип
эндогенный ингибитор ангиогенеза, был подтвержден как
многофункциональный противоопухолевый фактор. Сообщалось, что PEDF
ингибирует ангиогенез, индуцируя апоптоз ЭК и некоторых
типы опухолевых клеток (10,16). Полноразмерный PEDF имеет несколько
функциональные участки, а антиангиогенный функциональный фрагмент
расположены в остатках с 24 по 57 (известный как PEDF-34mer) (13,32). В текущем исследовании наши данные показали, что пептид P18,
сопоставление функционального фрагмента с остатками с 40 по 57 полноразмерного
PEDF проявляет антиангиогенную активность как in vitro, так и
в естественных условиях. По сравнению с полноразмерным PEDF пептид Р18
более стабилен и биосовместим, но обладает низкой антигенностью, что
указывает на потенциал пептида Р18 для применения в
антиангиогенная терапия у больных ГЦК.
В текущем исследовании пептид Р18 был подтвержден
оказывать антиангиогенное действие на HCC in vivo. CD31 это
в основном сосредоточены на границах между эндотелиальными клетками и
может рассматриваться как биомаркер ангиогенеза. Исследования
продемонстрировали, что в эндотелиальных клетках передача сигналов VE-кадгерина,
экспрессия и локализация коррелируют с проницаемостью сосудов
и опухолевый ангиогенез (33). ИГХ
окрашивание CD31 и VE-кадгерина в опухолевых тканях предполагает, что
пептид Р18 действует как мощный ингибитор ангиогенеза
в естественных условиях. Кроме того, изображения окрашивания IF показали, что
подавление фосфорилирования VEGFR2 может быть механизмом
с помощью которых пептид Р18 выполняет свои функции.
Ангиогенез зависит от дестабилизации ЭК,
диссоциация и миграция. При быстром росте солидных опухолей
гипоксическая среда внутри опухолевых тканей стимулирует
экспрессия матриксных металлопротеиназ и проангиогенных
цитокины, такие как MMP-2, MMP-9, VE-кадгерин и E-кадгерин
(34). ММП-2 и ММП-9 являются
доступен в HUVEC и отвечает за ремоделирование тканей
во время канцерогенеза, метастазирования опухоли и ангиогенеза (35,36).
Наше исследование показало, что дополнительный VEGF может усиливать секрецию
ММП-2 и ММП-9в HUVEC, что соответствует результатам
Анализы миграции и инвазии HUVEC. Однако HUVEC, обработанные
Пептид Р18 не проявляет ответа на дополнительный VEGF. Более того, мы
наблюдали аналогичные результаты по уровням экспрессии MMP-2 и MMP-9 в
Клетки HepG2, обработанные пептидом P18. Результаты выше конечно
предполагают, что пептид Р18 может выполнять свои антиангиогенные
функцию путем ингибирования биологической активности VEGF. Однако,
противоположные результаты были показаны в эффектах пептида Р18 на
Экспрессия Е-кадгерина в клетках HUVEC и HepG2. Повышение регуляции
E-кадгерина в клетках HepG2 отвечает за подавление
миграция клеток (37,38). Экспрессия Е-кадгерина в
HUVEC показали тенденцию к снижению. Мы считаем, что снижение
активность клеток после обработки пептидом Р18 может быть связана
к этому явлению. Являясь белком адгезии между клетками,
снижение экспрессии Е-кадгерина может привести к нестабильности и
распад сосудистых структур. Однако точный механизм
с помощью которого пептид Р18 подавлял экспрессию Е-кадгерина в
HUVEC нуждается в дальнейшем изучении, что станет следующим шагом нашего исследования.
исследовательская работа.
Подобно исходному PEDF и PEDF-34mer, P18
пептид ингибирует пролиферацию и индуцирует апоптоз HUVECs
in vitro и имеет IC50 320 нМ. Бкл-2
семейство состоит из многих эволюционно консервативных белков, которые могут
регулируют клеточный апоптоз посредством классического митохондриального
путь апоптоза (39). Бкл-2 это
антиапоптотический белок в этом семействе, тогда как Bax является
проапоптотический белок. Взаимодействия между способствующими смерти и
факторы, подавляющие смерть, регулируют динамическое равновесие, в котором
соотношение антиапоптотических и проапоптотических белков
контролирует клеточный апоптоз (40).
Согласно нашим результатам, HUVEC поддерживают низкий уровень апоптоза.
при определенной концентрации VEGF (8 нг/мл). Пептид Р18
значительно увеличивает скорость апоптоза, сопровождающегося
снижение как экспрессии Bcl-2, так и соотношения
Бкл-2/Вах. Однако, по нашим данным, лечение
Пептид Р18 не изменяет скорость апоптоза клеток HepG2.
На основании этих результатов мы предполагаем, что P18
пептид ингибирует ангиогенез, блокируя ось VEGF/VEGFR2 и
индуцирующие апоптоз эндотелиальных клеток. VEGF, который вырабатывается
ряд клеток, включая эндотелиальные клетки, макрофаги и
различных типов опухолевых клеток, участвует в ангиогенезе,
выживаемость эндотелиальных клеток сосудов, пролиферация и сосудистая
проходимость (3,20). Результаты миграции и
анализы образования трубок показали, что пептид P18 может ингибировать ECs
миграция и ангиогенная способность, индуцированная обработкой VEGF. К
дальнейшее объяснение этих явлений, фосфорилирование VEGFR2
в HUVECs. Как основной рецептор VEGF, VEGFR2-индуцированный
сигнализация необходима для выполнения VEGF-стимулированных
выживаемость, миграция ЭК и ангиогенез в развитии
опухолей (18,22). Фосфорилирование VEGFR2 приводит к
активация нижестоящих сигнальных путей, в том числе
Пути MAPK/Erk и PI3K/Akt (22,26).
По нашим данным, снижение уровня фосфорилирования
VEGFR2 в HUVEC наблюдали после культивирования в бессывороточной среде.
среде в течение 24 ч, и эти изменения быстро обратились в
наличие сыворотки или дополнительного VEGF. Однако это обращение было
отменяется при одновременном добавлении пептида Р18 к
средний. Такой же результат наблюдался и при добавлении SU1498.
вместо пептида Р18. Мы отметили, что лечение P18
пептид также подавлял уровни фосфорилирования PI3K и
Akt индуцируется VEGF, тогда как соотношение p-PI3K/PI3K и p-Akt/Akt
может повышаться при лечении ИФР-1, активатором
Путь PI3K/Akt. Кроме того, мы исследовали вариации в
экспрессия Bcl-2/Bax среди клеток HUVEC, обработанных пептидом P18,
контрольная группа и группа, получавшая IGF-1. С предыдущего
исследования показали, что снижение уровня Bcl-2/Bax приводит к
расщепление членов каспазы и инициирует каспазный каскад,
что приводит к активации и усилению клеточного апоптоза
ответов (41–43), мы обнаружили расщепленную каспазу-3 и
общая каспаза-3 как показатель для оценки апоптоза. Согласно с
результаты вестерн-блоттинга, мы подтвердили, что пептид Р18
подавлял экспрессию Bcl-2 и соотношение Bcl-2/Bax,
сопровождается усиленным расщеплением каспазы-3 синхронно с
ингибирование пути PI3K/Akt. Следовательно, лечение
Пептид Р18 был связан с усиленным митохондриально-опосредованным
апоптоз.
Таким образом, пептид Р18 оказывает свое
антиангиогенная биоактивность за счет ингибирования эндотелиальных клеток
жизнеспособность и индуцируют апоптоз. В то же время пептид Р18
подавляет жизнеспособность опухолевых клеток, подавляя миграцию клеток
вместо того, чтобы индуцировать апоптоз при ГЦК. VEGFR2 является основной мишенью
пептида Р18, действующего в ЭК. Путем модуляции VEGF/VEGFR2 и его
индуцирует сигнальный путь PI3K/Akt, пептид P18 усиливает
митохондриально-опосредованный апоптоз в ЭК и подавляет
образования в опухолевых тканях. Эта антиангиогенная активность
Пептид Р18 предполагает, что он может быть потенциальным агентом для
лечение ГЦК.
Благодарности
Это исследование было поддержано Национальным крупным Программа исследований и разработок Китая (2016YFC0106004), Планирование развития науки и технологий провинции Шаньдун, Китай (2015GGB14168) и естествознание провинции Шаньдун Фонд, Китай (ZR2015HL080).
Каталожные номера
1 | Carmeliet P: Ангиогенез в жизни, болезнь и медицина. Природа. 438: 932–936. 2005. Просмотр статьи : Google Scholar : PubMed/NCBI |
2 | Ханахан Д. |
3 | Schlieve CR, Mojica SG, Holoyda KA, Hou X, Fowler KL и Grikscheit TC: Сосудистый эндотелиальный фактор роста (VEGF) биодоступность регулирует ангиогенез и кишечный ствол и пролиферация клеток-предшественников во время постнатальной тонкой кишки разработка. ПЛОС Один. 11:e01513962016. Просмотр статьи : Google Scholar : PubMed/NCBI |
4 | Li Y, Turpin CP и Wang S: Роль тромбоспондин 1 при заболеваниях печени. Гепатол рез. 4 августа 2016 г. (Epub перед печатью). |
5 | Zhu AX, Duda DG, Sahani DV и Jain RK:
HCC и ангиогенез: возможные цели и будущие направления. |
6 | Атта М.М., Атта Х.М., Гад М.А., Рашед Л.А., Саид EM, Hassanien Sel-S и Kaseb AO: Клиническое значение сосудистой эндотелиальный фактор роста в гепатоцеллюлярной ткани, связанной с гепатитом С карцинома у египетских пациентов. J Гепатоцеллюлярная карцинома. 3:19–24. 2016. Просмотр статьи: Google Scholar: PubMed/NCBI |
7 | Welker MW и Trojan J: Антиангиогенез в лечении гепатоцеллюлярной карциномы: текущие данные и будущее перспективы. Мир J Гастроэнтерол. 17:3075–3081. 2011. PubMed/NCBI |
8 | Эделин Дж., Буше Э., Роллан Ю., Волеон
E, Pracht M, Perrin C, Le Roux C и Raoul JL: Сравнение опухолей
ответ по критериям оценки ответа при солидных опухолях (RECIST)
и модифицированный RECIST у пациентов, получавших сорафениб для
гепатоцеллюлярная карцинома. |
9 | Доусон Д.В., Вольперт О.В., Гиллис П., Кроуфорд SE, Xu H, Benedict W и Bouck NP: пигментный эпителий. фактор: мощный ингибитор ангиогенеза. Наука. 285: 245–248. 1999. Просмотр статьи : Google Scholar : PubMed/NCBI |
10 | He X, Cheng R, Benyajati S и Ma JX: PEDF и его роль в физиологических и патологических состояниях: Значение при диабетических и индуцированных гипоксией ангиогенных заболеваниях. Clin Sci (Лондон). 128:805–823. 2015. Просмотр статьи: Google Scholar: PubMed/NCBI |
11 | Belkacemi L и Zhang SX: Противоопухолевый
эффекты фактора пигментного эпителия (PEDF): значение
для терапии рака. Мини обзор. J Exp Clin Cancer Res. |
12 | Smith ND, Schulze-Hoepfner FT, Veliceasa D, Filleur S, Shareef S, Huang L, Huang XM и Volpert OV: Пигмент эпителиальный фактор и интерлейкин-6 контролируют простату нейроэндокринная дифференцировка по механизму прямой связи. Дж Урол. 179: 2427–2434. 2008. Просмотр статьи: Google Scholar: PubMed/NCBI . |
13 | Филлер С., Волц К., Нелиус Т., Мирочник Ю., Хуан Х., Зайчук Т.А., Аймерич М.С., Бесерра С.П., Яп Р., Величеаса Д., и др.: Два функциональных эпитопа фактора пигментного эпителия. блокируют ангиогенез и индуцируют дифференцировку при раке предстательной железы. Рак Рез. 65:5144–5152. 2005. Просмотр статьи : Google Scholar : PubMed/NCBI |
14 | Он СС, Ши ХС, Инь Т, Ли YX, Луо ST, Ву
QJ, Lu L, Wei YQ и Yang L: AAV-опосредованный перенос генов человека
Фактор пигментного эпителия ингибирует карциному легких Льюис
рост у мышей. |
15 | Broadhead ML, Dass CR и Choong PF: Системно вводимый PEDF против первичного и вторичного опухоли в клинически значимой модели остеосаркомы. Бр Дж Рак. 105: 1503–1511. 2011. Просмотр статьи : Google Scholar : PubMed/NCBI |
16 | Хасэ Р., Миямото М., Уэхара Х., Кадоя М., Эбихара Ю, Мураками Ю, Такахаши Р, Мега С, Ли Л, Шичинохе Т и др. al: Генная терапия фактора, полученного из пигментного эпителия, ингибирует человеческий рак поджелудочной железы у мышей. Клин Рак Рез. 11:8737–8744. 2005. Просмотр статьи : Академия Google : PubMed/NCBI |
17 | Мирочник Ю., Аврора А., Шульце-Хёпфнер
FT, Deabes A, Shifrin V, Beckmann R, Polsky C и Volpert OV: Short
пептид, полученный из пигментного эпителиального фактора, ингибирует
ангиогенез и рост опухоли. |
18 | Hicklin DJ и Ellis LM: Роль путь фактора роста эндотелия сосудов в росте опухоли и ангиогенез. Дж. Клин Онкол. 23:1011–1027. 2005. Просмотр статьи: Google Scholar: PubMed/NCBI . |
19 | Жсебик Б., Симонович К., Салех Ю., Зиолковски П., Бронович А. и Вереб Г.: Фотодинамическая терапия в сочетании с ингибитором цистеинпротеиназы синергически снижают выработку VEGF и способствуют некрозу опухоли у крыс рак молочной железы. Селл Пролиф. 40:38–49. 2007. Просмотр статьи: Google Scholar: PubMed/NCBI . |
20 | Каземи М., Каррер А., Моймас С., Зандона Л.,
Буссани Р., Касагранда Б., Пальмизано С., Прелацци П., Джакка М. |
21 | Фреззетти Д., Галло М., Рома К., Д’Алессио А., Майелло М.Р., Бевилаква С., Норманно Н. и Де Лука А.: Сосудистые эндотелиальный фактор роста а регулирует секрецию различных Ангиогенные факторы в клетках рака легкого. J Cell Physiol. 231: 1514–1521. 2016. Просмотр статьи : Google Scholar : PubMed/NCBI |
22 | Олссон А.К., Димберг А., Крюгер Дж. и Claesson-Welsh L: передача сигналов рецептора VEGF – в контроле сосудистой функция. Nat Rev Mol Cell Biol. 7: 359–371. 2006. Просмотр статьи: Google Scholar: PubMed/NCBI . |
23 | Robinson CJ и Stringer SE: сращивание
варианты фактора роста эндотелия сосудов (VEGF) и их
рецепторы. |
24 | Domigan CK, Ziyad S и Iruela-Arispe ML: Канонический и неканонический фактор роста эндотелия сосудов пути: новые разработки в области биологии и передачи сигналов. Артериосклеры Тромб Васк Биол. 35:30–39. 2015. Просмотр статьи: Google Scholar: PubMed/NCBI |
25 | Смит Н.Р., Бейкер Д., Джеймс Н.Х., Рэтклифф К., Дженкинс М., Эштон С.Э., Спроат Г., Суонн Р., Грей Н., Райан А. и др.: Рецепторы сосудистого эндотелиального фактора роста VEGFR-2 и VEGFR-3 локализуются преимущественно в сосудистой сети первичных солидных тканей человека. раки. Клин Рак Рез. 16:3548–3561. 2010. Просмотр статьи : Google Scholar : PubMed/NCBI |
26 | Holmqvist K, Cross MJ, Rolny C, Hägerkvist
R, Rahimi N, Matsumoto T, Claesson-Welsh L и Welsh M: Адаптер
белок shb связывается с тирозином 1175 при росте эндотелия сосудов
фактор (VEGF) рецептор-2 и регулирует VEGF-зависимую клеточную
миграция. |
27 | Джонстон Э.К., Фрэнсис М.К. и Неппер Дж.Э.: Рекомбинантный фактор пигментного эпителия PEDF связывает сосудистые рецепторы эндотелиального фактора роста 1 и 2. In Vitro Cell Dev Biol Аним. 51:730–738. 2015. Просмотр статьи : Google Scholar : PubMed/NCBI |
28 | Ву Л.Ф., Е. Ю. К., Хуан Г. Ю., Ли Х. Б., Ли Г. П., Пу ZJ, Wei BL и Feng JL: Участие стресса эндоплазматического ретикулума в индуцированном аденозином апоптозе клеток HepG2 гепатомы человека. Онкол Отчет 26:73–79. 2011. PubMed/NCBI . |
29 | Донато МТТЛ, Толоса Л. и Гомес-Лехон М.Х.:
Культура и функциональная характеристика гепатомы человека HepG2
клетки. |
30 | Киселевская Дж., Лигеза Дж. и Кляйн А.: действие ингибиторов тирозинкиназы, тирфостинов: AG1024 и SU1498 на аутокринный рост клеток рака предстательной железы (DU145). Фолиа Гистохим Цитобиол. 46:185–191. 2008. Просмотр статьи: Google Scholar: PubMed/NCBI . |
31 | Лаурино Л., Ван ХХ, де ла Уссе Б.А., Соса Л., Дюпраз С., Касерес А., Пфеннингер К.Х. и Кирога С.: PI3K Активация ИФР-1 необходима для регуляции мембранных расширение конуса роста нерва. Дж. Клеточные науки. 118:3653–3662. 2005. Просмотр статьи : Google Scholar : PubMed/NCBI |
32 | Гонг Кью, Цю С, Ли С, Ма Ю, Чен М, Яо Ю,
Che D, Feng J, Cai W, Ma J и др. |
33 | Кун Б.Г., Байенс Н., Хан Дж., Будата М., Росс Т.Д., Фанг Дж.С., Юн С., Томас Дж.Л. и Шварц М.А.: Внутримембранный связывание VE-кадгерина с VEGFR2 и VEGFR3 собирает эндотелиальный механосенсорный комплекс. Джей Селл Биол. 208:975–986. 2015. Просмотр статьи : Академия Google : PubMed/NCBI |
34 | Ляо Д. и Джонсон Р.С.: Гипоксия: ключ регулятор ангиогенеза при раке. Метастаз рака Rev. 26:281–290. 2007. Просмотр статьи: Google Scholar: PubMed/NCBI . |
35 | Штернлихт М.Д. и Верб З.: Как матрица
металлопротеиназы регулируют поведение клеток. |
36 | Li H, Daculsi R, Bareille R, Bourget C и Amedee J: uPA и MMP-2 были вовлечены в самособирающуюся сеть формирование в двумерной модели совместной культуры костного мозга стромальные клетки и эндотелиальные клетки. Джей Селл Биохим. 114: 650–657. 2013. Просмотр статьи: Google Scholar: PubMed/NCBI . |
37 | Чжао И, Пэн С, Цзя С, Сюй Ф, Сюй И и Дай C: Armc8 регулирует инвазивную способность гепатоцеллюлярной карциномы. через комплекс Е-кадгерин/катенин. Опухоль биол. 37:11219–11224. 2016. Просмотр статьи: Google Scholar: PubMed/NCBI |
38 | Yu AQ, Ding Y, Li CL, Yang Y, Yan SR и
Li DS: вызванное TALEN нарушение экспрессии Nanog приводит к
снижение пролиферации, инвазивности и миграции, увеличение
химиочувствительность и реверсирование ЭМП в клетках HepG2. |
39 | Кори С., Хуанг Д.К. и Адамс Дж.М.: Bcl-2 семья: роль в выживании клеток и онкогенезе. Онкоген. 22:8590–8607. 2003. Просмотр статьи: Google Scholar: PubMed/NCBI . |
40 | Кори С. и Адамс Дж. М.: Семейство Bcl2: Регуляторы клеточного переключателя жизни и смерти. Нат Рев Рак. 2: 647–656. 2002. Посмотреть Статья : Google Scholar : PubMed/NCBI |
41 | Li LY, Luo X и Wang X: эндонуклеаза G апоптотическая ДНКаза при выходе из митохондрий. Природа. 412: 95–99. 2001. Просмотр статьи: Google Scholar: PubMed/NCBI |
42 | Тянь X, Ши Ю, Лю Н, Ян Ю, Ли Т, Хуа П
и Лю Б. |
43 | Цао И, Цзян Зи, Цзэн Зи, Лю И, Гу И, Цзи И, Чжао И и Ли И: подавление Bcl-2 ослабляет индуцированную гипоксией резистентность к апоптозу в эндотелиальных клетках легочных микрососудов. Апоптоз. 21:69–84. 2016. Просмотр статьи: Google Scholar: PubMed/NCBI |
Регрессия созревания клубочков определяет популяцию нефронов у нормальных мышей
Main
Во время развития органов полная регрессия одной структуры часто сопровождается появлением и ростом других структур (1). При развитии почки появляются пронефрос и мезонефрос, которые затем регрессируют до структурного развития метанефроса, который развивается в зрелую почку (2). Формирование с последующей частичной регрессией также происходит во время нормального развития (3). Уменьшение количества нефронов в процессе развития in utero был связан с рядом заболеваний (4,5,6), ситуацию, которая может быть компенсирована вмешательством во время постнатального созревания (7,8,9,10). Во время нашего экспериментального исследования развития почек мышей мы неожиданно обнаружили, что количество клубочков у мышей вскоре после рождения уменьшилось. Мы предположили, что регуляторная регрессия избыточной клубочковой сосудистой сети является механизмом поддержания гомеостаза числа клубочков. В этом исследовании мы исследовали, происходит ли регрессия после нормального гломерулогенеза.
Развитие почек у мышей в постнатальном периоде происходит параллельно развитию поздних эмбриональных стадий у людей, когда большинство клубочков уже сформировано. Таким образом, процесс постнатального созревания у мышей до возраста 4 недель считается моделью развития почек эмбриона человека. Чтобы воспроизводимо оценить количество клубочков у постнатальных мышей, мы использовали беспристрастный стереологический метод подсчета проб физического фракционера с подсчетом диссекторов (метод фракционирования/диссектора) (11,12). Кроме того, мы изучили применимость метода Weibel-Gomez для оценки числа клубочков, который обычно используется в зрелых почках (13).
Результаты
Идентификация клубочков в постнатальных почках мышей для подсчета
Поскольку на ранних стадиях развития клубочков распознаются только типичные запятые или S-образные клубочки, мы использовали маркеры дифференцировки подоцитов для облегчения идентификации клубочков. В постнатальный день (P) 3 почки положительные сигналы опухоли Вильмса 1 не были локализованы исключительно в клубочках, а поверхностно развивающиеся клубочки не были легко различимы при окрашивании синаптоподином и периодической кислотой-Шиффом (PAS) ( Рисунок 1 ). Кроме того, очертания возможных клубочков были недостаточно четкими для определения площади клубочков, требуемой методом Weibel-Gomez. Напротив, окрашивание синаптоподином легко идентифицировало клубочки на Р7. Почки мышей старше P10 имели клубочки, которые легко распознавались окрашиванием PAS. Таким образом, только почки от мышей старше P7, имеющие четко идентифицируемые клубочки, были исследованы на количество клубочков методами Weibel-Gomez и методами фракционирования/диссектора.
Идентификация клубочков в постнатальных почках мышей. Соседние срезы окрашивали на PAS, WT1 и синаптоподин. На P3 поверхностные клубочки не могли быть идентифицированы исключительно тремя окрашиваниями; однако их можно было различить по синаптоподину на P7. Все клубочки можно было легко распознать с помощью PAS, начиная с P10. Черные стрелки (средний столбец) указывают клубочек, который можно идентифицировать с помощью окрашивания синаптоподином и WT1, но не с помощью PAS. Р, послеродовой день; PAS, периодическая кислота-Шифф; Син, синаптоподин; WT1, опухоль Вильмса 1. Полоса = 50 мкм.
Изображение в натуральную величину
Оценка клубочкового числа по методу Weibel-Gomez
Сначала мы исследовали вариабельность клубочкового числа между левой и правой почками у одной и той же мыши по методу Weibel-Gomez (13,14). Почки от трех до пяти мышей C57BL/6J исследовали в каждый момент времени с P7 до P28. Не было никаких существенных различий между левой и правой почками в любой момент времени, а количество клубочков в левой почке достоверно коррелировало с таковым в правой почке (9).0443 R 2 = 0,320, P < 0,001). Таким образом, мы использовали количество клубочков в правой почке, полученное методом Вейбеля-Гомеса, и сравнили с количеством клубочков, полученным методом фракционирования/диссектора из левой почки.
Межэкспертная изменчивость проверялась путем случайной выборки одной почки в каждый момент времени с P7 по P28. Корреляция в семи временных точках была значимой как для средней площади клубочков ( R 2 = 0,949, P < 0,001) и клубочкового числа ( R 2 = 0,507, P < 0,05). Мы оценили необходимость исследования всех клубочков на срезе для определения средней площади клубочков (1). С этой целью 25% или 50% всех клубочков, выбранных случайным образом, сравнивали со всеми клубочками на срезе. Корреляционные анализы показали R 2 = 0,968 для доли 25% и R 2 = 0,942 при оценке 50% клубочков, оба P <0,001. Таким образом, на центральном поперечном срезе почки мыши отбор 40 случайно выбранных клубочков или клубочков из непрерывных полей, которые включают как поверхностные, так и глубокие клубочки, достаточен для получения согласованных значений числа клубочков по методу Weibel-Gomez.
Плотность почек, коэффициент усадки и коэффициент распределения клубочков по размерам в постнатальных почках мышей
Первоначально мы использовали установленные коэффициенты плотности почек (1,04 г/см 3 ) и усадки (1,08), которые обычно используются для взрослых крыс (14). Однако эти коэффициенты дали гораздо большее количество клубочков, чем те, которые были измерены методом фракционирования/диссектора во все моменты времени. Чтобы определить более подходящее значение для коэффициентов, применяемых в формуле Вейбеля-Гомеса для оценки почек мышей, необходимо точно определить объем почек при сборе и после фиксации и обработки. Мы использовали микрокомпьютерную томографию и технологию трехмерной реконструкции изображения, чтобы получить объем почки мыши ( Рисунок 2 ). Наши результаты показали, что общее уменьшение почки мыши было намного выше, чем у крыс. Плотность почек и фракция усадки в развивающихся почках мышей от P7 до P14 были значительно выше, чем в почках после P18. После П18 плотность и коэффициент усадки стабилизировались и составили 0,968 (0,913–1,023) г/см 3 для плотности почек и 1,395 (1,373–1,416) для усадки ( Стол 1 ). Чтобы упростить уравнение, используемое для метода Вейбеля-Гомеса, мы ввели поправочный коэффициент f , который представляет собой массу свежесобранной почки, деленную на объем почки после фиксации и обработки (подробно в разделе «Материалы и методы»). Для нормальных почек после P18 мы нашли среднее значение f , равное 2,62.
Репрезентативные микрокомпьютерные томографические изображения почки мыши. ( a ) Сканирование изображений соседних слоев показало четкое отделение ткани от периферической, а полуавтоматическое оконтуривание четко очертило образец почки. Цифры на подпанелях указывают порядковые номера срезов образца. ( b ) Репрезентативное увеличенное изображение для контуринга. ( c ) Трехмерная реконструкция изображения почки из отсканированных слоев, используемая для расчета общего объема почки. Бар = 1 мм.
Полноразмерное изображение
Таблица 1 Коэффициенты, разработанные для подсчета клубочков по формуле Вейбеля-Гомеза у развивающихся мышейПолноразмерная таблица
Заметная разница в зрелости и размере клубочков характеризует поверхностные и глубокие клубочки в постнатальных почках мышей. Однако мы обнаружили, что коэффициент распределения клубочков по размерам κ (15, 16), рассчитанный на основе измеренных профилей диаметра срезов клубочков, был относительно постоянным у мышей разного возраста и был сравним с таковым, полученным в исследованиях почек взрослых крыс (14) и образцах почек человека (16).
Сравнение клубочкового числа, рассчитанного по методу Weibel-Gomez и измеренного методом фракционирования/диссектора
Используя исчерпывающий метод фракционирования/диссектора, мы обнаружили, что клубочковое число было максимальным на P7. За этим последовало сокращение клубочков до P18. Различия в количестве клубочков на П18, П21, П25 и П28 по сравнению с П7 были статистически значимыми ( Таблица 2 , Рисунок 3 ).
Таблица 2 Клубочковое число, определенное методом Вейбеля-Гомеса и исчерпывающим методом фракционирования/диссектора у развивающихся мышейПолноразмерная таблица
Рисунок 3 ) исчерпывающий метод фракционирования/диссектора у нормально развивающихся мышей. В то время как метод Вейбеля-Гомеса давал непрерывное увеличение числа клубочков у постнатальных мышей, метод фракционирования/диссектора выявил значительное уменьшение после 14-го дня. Данные представлены как среднее значение ± стандартная ошибка среднего. * P < 0,05 по сравнению с 7-м днем постнатального периода. метод фракционирования/диссектора после P18. Однако метод Вейбеля-Гомеза дал гораздо меньшее количество клубочков, чем метод фракционирования/диссектора до P18 (
Стол 2 , Рисунок 4 ). Рисунок 4 Попарное сравнение числа клубочков, оцененных методом Weibel-Gomez и методом ракционатора/диссектора у одной и той же мыши. В развивающихся почках мышей количество клубочков, подсчитанное по методу Вейбеля-Гомеса, было значительно ниже, чем по методу фракционирования/диссектора на 7, 10 и 14 постнатальные дни (все P <0,05), но аналогично после 18 постнатального дня. Послеродовые дни ( a ) 7, ( b ) 10, ( c ) 14, ( d ) 18, ( e ) 21 и ( f ) 25–28. F/D, метод фракционирования/диссектора; W–G, метод Вейбеля–Гомеса.
Изображение полного размера
Гломерулярная регрессия в постнатальных почках мышей
Для изучения потенциальных механизмов, ответственных за уменьшение числа клубочков после П7, мы оценили факторы, участвующие в сосудистой регрессии, ангиопоэтин 2 (ANGPT2) (17,18) и тромбоспондин. 1 (THBS1) (19,20). Иммуноокрашивание ANGPT2 и THBS1 присутствовало в клубочках, проксимальных канальцах и интерстиции в постнатальных почках мышей. Рисунок 5 ). В клубочках ANGPT2- или THBS1-положительные клетки включали подоциты, эндотелиальные клетки, клетки мезангиальной области и, вероятно, перициты. Как поверхностные, так и глубокие клубочки экспрессируют ANGPT2 и THBS1, причем более выраженная экспрессия наблюдается в поверхностной коре. Гломерулярная экспрессия ANGPT2 достигала пика на P14 и была значительно выше, чем на P7, P10, P25 и P28. У мышей также была пиковая экспрессия THBS1 на P14, которая была значительно выше, чем на P7, P25 и P28. Рисунок 5 ). Всплеск экспрессии ANGPT2 и THBS1 свидетельствует о нормальном регуляторном контроле роста клубочков и регрессии избыточных капилляров клубочков в течение позднего периода созревания после завершения гломерулогенеза.
Гломерулярная экспрессия ангиопоэтина 2 (ANGPT2) и THBS1 в нормально развивающихся почках мышей. В клубочках ( a ) ANGPT2 и ( b ) THBS1 экспрессируются в подоцитах, эндотелиальных клетках, мезангиальных клетках и предполагаемых перицитах. Количество ANGPT2- или THBS1-положительных клубочков достигало пика на P14, что было значительно больше, чем в большинстве других изученных моментов времени (9).1344 c и d ). Данные представлены как среднее ± SEM. * P <0,05 по сравнению с P14. Р, послеродовой день; THBS1, тромбоспондин 1. Бар = 20 мкм.
Изображение в полный размер
Для дальнейшего подтверждения результатов динамических изменений этих маркеров регрессии у различных линий мышей дополнительные образцы почек мышей BALB/c и 129/SvJ были исследованы с помощью иммуногистохимии на P7, P14 и P21. Значительная активация ANGPT2 и THBS1 происходила на P14 как в BALB/c, так и в 129/SvJ мыши ( Рисунок 6 ), напоминающие результаты у мышей C57BL/6J.
Гломерулярная экспрессия ангиопоэтина 2 (ANGPT2) и тромбоспондина 1 у ( a ) нормально развивающихся мышей BALB/c и ( b ) 129/SvJ. Данные представлены как среднее ± SEM. * P < 0,05 по сравнению с 7-м днем после рождения. † P < 0,05 по сравнению с 21-м днем после рождения. THBS1, тромбоспондин 1.
Изображение в натуральную величину
Обсуждение
Используя метод подсчета фракционирования/диссектора, это исследование показывает, что максимальное число клубочков происходит на P7, за которым следует снижение примерно на 20% до P18 у нормально развивающихся мышей. Эти изменения дополняются повышенной экспрессией маркеров сосудистой регрессии ANGPT2 и THBS1, которые увеличиваются и достигают пика в созревающих клубочках на P14. Наше исследование подтверждает использование метода Вейбеля-Гомеса после P18 для оценки общего числа клубочков в нормальной почке мыши с обоснованием коэффициентов, но сужает его применение в незрелых почках.
Для изучения клубочкового числа использовалось несколько методологий, включая метод фракционирования/диссектора, уравнение Вейбеля-Гомеса и метод диссоциации (21). Недавние технологические достижения привели к врожденным аномалиям почек в отношении размера и числа нефронов, что требует точного, воспроизводимого и практичного подхода к определению числа клубочков на стадиях развития. Метод фракционирования/диссектора является беспристрастным для подсчета количества клубочков в почках на любой стадии развития (11,12). Однако эта методология золотого стандарта требует истощения ткани. Этот метод также трудоемкий и трудоемкий. Метод диссоциации также включает в себя истощение ткани и имеет ограничение высокой изменчивости (21). Метод Вейбеля-Гомеса был разработан в начале 19 века. 60 с и использовался для оценки количества нефронов во всей почке, предположительно, на репрезентативном двухмерном срезе почки (13). Однако значения коэффициентов, используемых в формуле Вейбеля-Гомеса, не были проверены на почках мышей. Кроме того, эти коэффициенты не были проверены на незрелых почках мышей или даже крыс. Мы показали, что плотность почек и фракция усадки, переменные, определяющие коэффициенты в формуле Weibel-Gomez, становятся постоянными только после P18. Несмотря на высокую воспроизводимость метода Weibel-Gomez между исследователями, а также между двумя почками одного и того же животного, наши результаты показывают, что метод Weibel-Gomez занижает количество клубочков в незрелой почке. Мы считаем, что этот метод непригоден для оценки клубочковой популяции у нормальных мышей до P18. Этот недоучет в первую очередь связан со значительно меньшим размером незрелых поверхностных клубочков, что приводит к систематической ошибке отбора. Полученные данные свидетельствуют о необходимости осторожности при использовании некоторых морфометрических методов, основанных на измерениях, полученных из одного среза, для профилирования незрелой ткани, например, при расчете плотности структуры для сравнения нормальных и больных почек. Эти ограничения влияют на исследования, направленные на экстраполяцию наблюдаемых аномалий развития для прогнозирования будущих заболеваний.
Сообщается, что гломерулогенез заканчивается на P3 у мышей (22), тогда как недавнее исследование развивающихся почек крыс показало продолжающееся увеличение числа клубочков до P8 (12). Хотя неизвестно, как обструкция мочеточника влияет на развитие клубочков, у крыс с односторонней обструкцией мочеточника на P1 или P14, высвобождаемой через 5 дней, наблюдалась одинаковая степень снижения количества нефронов во взрослом возрасте (23). Другие исследования у новорожденных крыс с унинефрэктомией показали различные возможности роста клубочков в оставшейся почке в зависимости от времени операции (7,24). В этом исследовании мы стремились определить время для возможной компенсации и/или вмешательства для достижения нормального набора клубочков. На результаты, полученные методом фракционирования/диссектора, не влияет распределение клубочков в почке, форма, объем или деформация в результате обработки ткани. Клубочковое число, оцененное с помощью этого золотого стандарта, указывает на то, что пик клубочкового числа приходится на P7. Это наблюдение согласуется с предыдущими выводами на крысах (7,12), видах, время развития которых сходно с мышами. Будущие исследования потребуются, чтобы определить биологическую важность этого динамического изменения числа клубочков во время созревания в патофизиологических условиях.
Уменьшение числа клубочков дополняется повышенной экспрессией клубочков ANGPT2 и THBS1, двух хорошо известных естественных ингибиторов ангиогенеза. ANGPT2 противодействует эффектам васкулогенеза и ангиогенеза фактора роста эндотелия сосудов и ангиопоэтина 1, способствуя регрессии или дегенерации сосудов (17,18). В развивающихся клубочках повышенная регуляция ANGPT2 была обнаружена на подоцитах, эндотелиальных клетках и мезангиальных клетках (25,26). Экспрессия мРНК ANGPT2 в развивающихся почках мышей достигает пика на 2-й неделе постнатального периода (27). В моделях взрослых животных со сверхэкспрессией ANGPT2 в подоцитах, гломерулонефритом и диабетической нефропатией повышение уровня ANGPT2 связано с протеинурией, потерей гломерулярных капилляров и гломерулосклерозом (17,28). THBS1 — еще один эндогенный мощный ингибитор ангиогенеза, а также проапоптотический фактор (19).). Было показано, что миметические пептиды THBS1 регрессируют установленную злокачественность, индуцируя апоптоз в незрелых эндотелиальных клетках (20). Также сообщалось о гломерулярной и почечной тубулоинтерстициальной экспрессии THBS1 в развивающихся и больных почках (29,30). В сетчатке мышей P28 новообразованные сосуды регрессировали у дикого типа, в то время как плотность сосудов увеличивалась у мышей с дефицитом THBS1 (31). В соответствии с этими наблюдениями наши данные о том, что ANGPT2 и THBS1 значительно активируются на P14 и преимущественно в поверхностных незрелых клубочках, предполагают контроль созревания избыточной клубочковой сосудистой сети и физиологический ответ на функциональные изменения этих клубочков на этой стадии развития. Кроме того, увеличение ANGPT2 и THBS1 на P14 также было обнаружено у других нормальных линий мышей, а именно BALB/c и 129./SvJ мыши.
Таким образом, наше исследование показывает, что пик клубочкового числа приходится на P7 у мышей, за которым следует примерно 20% снижение полного созревания числа нефронов. Эти изменения сопровождаются пиковой экспрессией маркеров сосудистой регрессии ANGPT2 и THBS1 в клубочках на P14. Эти результаты предполагают, что время до P14 является окном для корректировки числа нефронов при нормальном развитии клубочков. Мы также показываем, что коэффициенты в широко используемой формуле Weibel-Gomez для подсчета числа клубочков должны быть скорректированы на стадии развития.
Материалы и методы
Животные
Исследовали самцов и самок мышей C57BL/6J (Лаборатория Джексона, Бар-Харбор, Мэн), если не указано иное. Развивающиеся мышиные почки собирали на P7, P10, P14, P18, P21, P25 и P28. Почки мышей в возрасте 12–18 недель служили в качестве зрелых контролей. Инбредных мышей BALB/c (первоначально из Harlan Laboratories, Prattville, AL) и мышей 129/SvJ (первоначально из лаборатории Джексона) на P7, P14 и P21 также использовали для изучения экспрессии маркеров сосудистой регрессии в почках. Все животные содержались в контролируемых условиях в помещениях для животных, сертифицированных Ассоциацией по оценке и аккредитации ухода за лабораторными животными, и получали стандартный корм и воду ad libitum , со щенками, выкармливаемыми матерями до возраста отъема. Протоколы для животных были одобрены Институциональным комитетом по уходу и использованию животных Университета Вандербильта.
Препарат тканей
Взвешивали левую и правую почки. Центральный поперечный срез почки использовали для оценки числа клубочков по методу Weibel-Gomez, тогда как всю почку использовали по методу фракционирования/диссектора. Образцы почек фиксировали в течение ночи в 4% параформальдегиде в фосфатно-солевом буфере и хранили в 70% этаноле до дальнейшей обработки (Excelsior ES Tissue Processor; Thermo, Waltham, MA), где образцы обезвоживали посредством шести смен этанола определенной степени очистки, очищали в ксилоле, пропитывают и заливают парафином.
Оценка плотности почки и усадки ткани с помощью микрокомпьютерной томографии
Для оценки плотности почки и усадки во время подготовки ткани использовали микрокомпьютерную томографию для определения объема свежесобранной почки ( V o ) и объем после фиксации и обработки ( V p ). Томографические изображения были получены в µCT40 (Scanco Medical AG, Брюттизеллен, Швейцария) с изотропным размером вокселя 36 мкм, при 45 кВ, 177 мкА, 250 проекций на поворот на 180° и времени интегрирования 150 мс. Используя программное обеспечение производителя, внешний край почки на каждом изображении среза был очерчен с использованием полуавтоматического порогового процесса обнаружения. Общий объем почки рассчитывали путем создания Z-стека двумерных контуров и прямого преобразования общего количества вокселей в трехмерной реконструкции в общий объем. Плотность почек ( ρ ) рассчитывали путем деления веса на объем свежей ткани. Коэффициент усадки ( δ ) рассчитывали как корень кубический из доли объема, т.е.
Подсчет клубочков по методу Вейбеля-Гомеса
Центральный фрагмент почки, срез толщиной 3 мкм, окрашивали PAS. При расчете общего числа клубочков ( N glom ) использовалось следующее уравнение, разработанное Вейбелем и Гомесом (13,14):
, где N – количество клубочков, которое можно подсчитать на срезе ткани, W почки, – масса свежесобранной почки, – общая площадь всех клубочков, A почка, – площадь почки κ — коэффициент распределения по размерам, β — константа формы, равная ≈ 1,382 для сферы (15), δ — коэффициент усадки ткани, ρ — плотность свежей почка. Вышеприведенное уравнение также может быть выражено как:
где средняя площадь клубочка.
Потому что W почки /(Δ 3 × ρ) равно V P , побочный продукт Δ 9 3 и ratifit 444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444444 гг. f , для которого требуется только однократное сканирование микрокомпьютерной томографии для V p . Таким образом, формула Вейбеля-Гомеса также может быть выражена как:
, где f — поправочный коэффициент, который можно определить и настроить для каждой отдельной лаборатории.
ImageJ (http://rsbweb.nih.gov/ij/) использовали для измерения площадей на срезах, окрашенных PAS. Площадь клубочков измеряли путем отслеживания внутренней границы пространства Боумена и области почек путем отслеживания контура среза почечной ткани, включая почечный сосочек. Диаметр (ϕ) каждого измеренного клубочка рассчитывали по площади клубочка на срезе, вычисляли среднее значение ϕ и среднее значение ϕ 3 . Затем рассчитывали коэффициент распределения размеров κ для каждой почки как (15,16). Количество клубочков исследовали два обученных исследователя.
Подсчет клубочков методом фракционирования/диссектора
Каждую почку тщательно разрезали на срезы толщиной 5 мкм. Каждая 20-я секция и соседняя с ней секция отбирались как пара, причем первая пара выбиралась случайным образом, обеспечивая долю выборки 1/20. Всего для каждой почки было получено 17–23 пары срезов, которые были окрашены PAS. Световой микроскоп (Nikon ECLIPSE E400; Nikon Instruments, Мелвилл, Нью-Йорк), оснащенный цифровой камерой (Zeiss AxioCam, Оберкохен, Германия), использовался для захвата изображений, на которые была установлена сетка, служащая физическим диссектором для облегчения исследования. всех полей. В паре проб клубочек считался, если он появлялся в поле пробы, но отсутствовал в соседнем срезе (правило появления-исчезновения). Также подсчитывали клубочки, присутствующие в соседнем отделе, но отсутствующие в первом отделе. Таким образом, согласно стереологическому принципу Кавальери (11,15), общее число клубочков в почке равно сумме подсчитанного числа клубочков из каждой пары образцов, умноженной на 20.
Иммуногистохимия
Срезы размером три микрометра окрашивали наборами Vectatain ABC (Vector Laboratories, Burlingame, CA) для кроликов на опухоль Вильмса 1, синаптоподин, ANGPT2 и THBS1. Вкратце, срезы нагревали в буфере цитрата натрия с концентрацией 0,01 моль/л (рН 6,0) и гасили эндогенную пероксидазу свежим раствором 0,3% пероксидазы водорода/метанола в течение 5 мин. После блокировки срезы инкубировали с кроличьим антителом против опухоли Вильмса 1 (Santa Cruz Biotechnology, Санта-Крус, Калифорния), мышиным антисинаптоподином (Progen, Heidelberg, Germany), мышиным антителом к ANGPT2 (Santa Cruz Biotechnology) или мышиным антителом к THBS1 ( Santa Cruz Biotechnology) во влажной камере при 4 °C в течение ночи с последующей инкубацией с биотин-конъюгированным вторичным антителом и комплексом авидин-биотин-пероксидаза. Положительные сигналы проявлялись в коричневом цвете диаминобензидином с гематоксилином в качестве контрастного красителя. Клубочки, положительные для ANGPT2 или THBS1, подсчитывали под линзой объектива × 40 и рассчитывали процент.
Статистический анализ
Результаты представлены как средние (95% доверительный интервал среднего). Односторонний ANOVA и непараметрический критерий Крускала-Уоллиса с последующим апостериорным многократным сравнительным тестом Тьюки и Данна использовались для проверки различий между различными временными точками. Корреляционный анализ проводили с помощью тестов Пирсона и Спирмена. Для сравнения данных, полученных двумя разными методами, использовали ранговый критерий Уилкоксона и парные t -тесты. Значение P <0,05 считалось значимым.
Заявление о финансовой поддержке
Это исследование было поддержано Американской кардиологической ассоциацией (грант 09BGIA2261364) и Национальным институтом диабета, болезней органов пищеварения и почек, Национальным институтом здравоохранения (гранты DK037868 и DK044757).